Les pores de fusion dépendants des SNARE sont généralement considérés comme des structures transitoires.
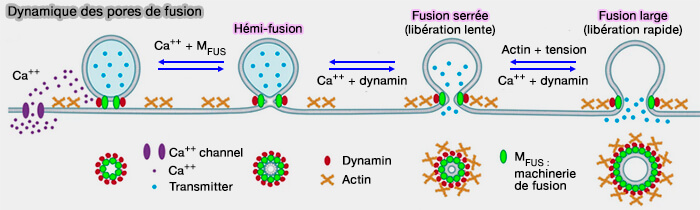
Les pores de fusion sont métastables, mais ils peuvent osciller (pore flickering) entre une fusion complète ou un retour à une hémifusion ou une fission.
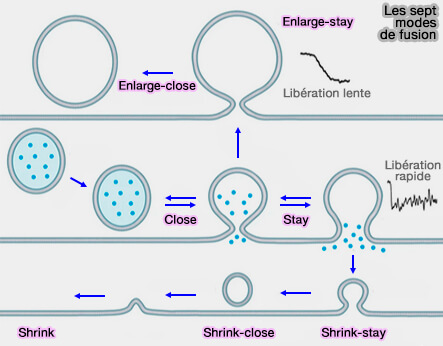
Dynamique des pores de fusion
C'est dans ce domaine que les connaissances ont le plus progressé ces dernières années, en particulier dans les vésicules synaptiques grâce à la microscopie STED (Stimulated Emission Depletion ou déplétion par émission stimulée).
Les conséquences de ces observations ont permis de redéfinir le kiss-and-run, souvent remis en question et considéré tout au plus comme un mode mineur de fusion/fission.
3. Les pores de fusion jusqu'à 490 nm peuvent rester ouverts ou se rétrécissent et se ferment en fonction du changement de taille du profil Ω.
à l'énergie libre liée à l'extension de son périmètre membranaire courbe, i.e. il faudrait la présence d'une force extérieure, comme la présence de pression osmotique ou une couche de protéines en croissance sur le col du pore de fusion,
à la création d'une zone membranaire en excès lors de l'expansion des pores (croissance du volume de la vésicule).
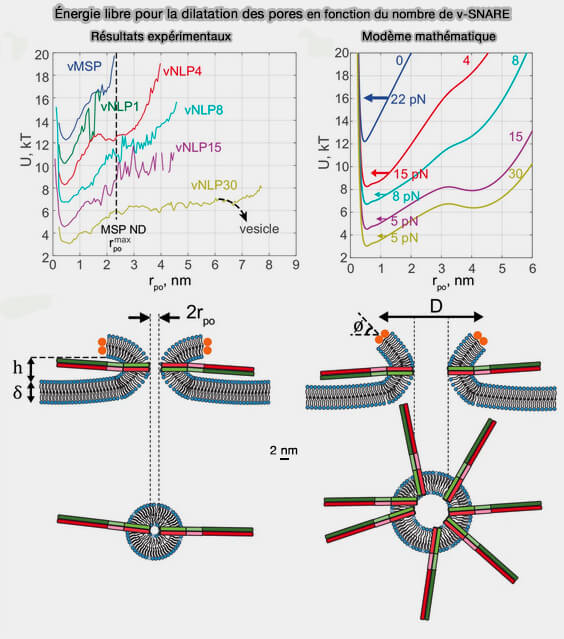
Énergie libre pour la dilatation des pores en fonction du nombre de v-SNARE
(Figure : vetopsy.fr d'après Wu et coll)
3. L'expansion du pore de fusion peut continuer jusqu'à ce que la vésicule s'aplatisse pour favoriser la libération, i.e. appelée fusion par effondrement complet (full-collapse fusion), mais, comme nous l'avons vu plus haut, ce phénomène est plutôt rare ( retrécissement versus effondrement complet).
Dynamique des pores de fusion
(Figure : vetopsy.fr d'après Wu et Chan)
2. Si l'influx calcique peut activer la dynamine pour permettre la constriction et la fermeture des pores, son blocage ou celui de la dynamine inhibe la constriction/fermeture des pores de fusion et augmente ainsi la taille initiale des pores ( fusion versus fission des pores).
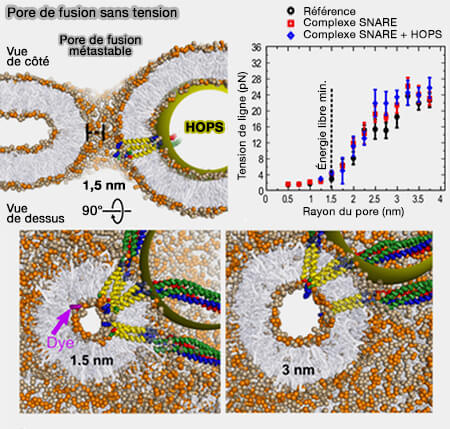
1. À des tailles de pores <1 nm, les répulsions d'hydratation domineraient l'énergie libre du pore. Cela suggère que le pore de fusion in vivo doit être soumis à une force de " compression " supplémentaire qui compense la répulsion de l'hydratation luminale dont la nature est floue à l'heure actuelle.
a. Il est possible que la lumière du pore soit influencée par des attractions électrostatiques.
En outre, elles inhiberaient la formation du diaphragme d'hémifusion, car l'hémifission diminuerait le nombre de groupes de têtes désormais favorables aux interactions à l'intérieur du pore.
b. Si on augmente de 5% les lipides des feuillets externes, cette asymétrie peut faciliter les pores de taille inférieure au nanomètre, en particulier lorsque les lipides de type PE (phosphatidyléthanolamine) tapissent en outre l'intérieur du pore.
Les vacuoles reliant les pores de fusion nanoscopiques stables pourraient donc s'expliquer par des différences de densité lipidique des feuillets et de leur composition en conséquence de l'amarrage vacuole-vacuole.
2. Un pore tapissé de plusieurs complexes SNARE affiche une plus grande stabilité contre l'hémifission, conformément aux observations expérimentales.
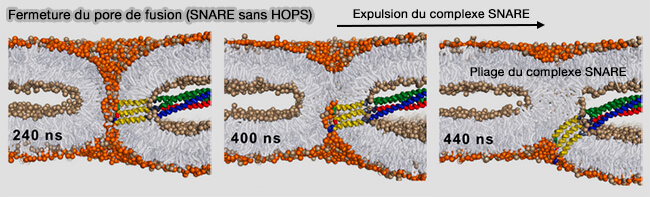
a. Un pore qui se ferme expulse activement les SNARE et les plie pour générer une contrainte.
Fermeture du pore de fusion (SNARE sans HOPS)
(Figure : vetopsy.fr d'après D'agostino et coll)
b. Ce mouvement devrait être entravé par la rigidité et la longueur du complexe SNARE, ainsi que par son attachement au complexe HOPS, qui est ancré dans les deux membranes.
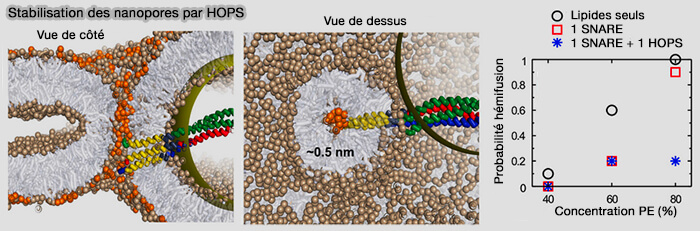
La répulsion stérique que HOPS impose au site de fusion réduit la courbure effective de l'intérieur du pore, ce qui, par analogie avec le rôle de la courbure dans la fusion membranaire, stabilise le pore en s'opposant à la formation d'une tige d'hémifission.
Un seul complexe SNARE avec HOPS réduit la probabilité de fermeture des pores jusqu'à 80%.
Stabilisation des nanopores par le complexe HOPS
(Figure : vetopsy.fr d'après D'agostino et coll)
Ce contact qui n'est pas observé en présence d'un intermédiaire d'hémifusion formé ou d'un pore de fusion soulève la question de savoir comment ces états ancrés sont mobilisés pour la fusion.
Les bicouches empilées sont séparées de 1,1 et 1,3 nm, distance supérieure à celle la fusion membranaire qui nécessite une distance inférieure à 1 nm afin de former un pont lipidique crucial.
L'adhérence s'oppose très probablement à la fois à la formation et à l'expansion du pore de fusion ainsi qu'à la formation d'intermédiaires d'hémifusion, car cela réduirait la quantité de surface de contact membrane-membrane favorable.
b. L'adhérence pourrait aider à surmonter les affrontements stériques et aider à la fois la fermeture éclair et la structuration du complexe SNARE en fournissant une force motrice supplémentaire et peut-être essentielle pour rapprocher suffisamment les membranes.
Petites versus grandes vésicules
Il n'est pas évident que les paramètres pertinents pour une fusion efficace se chevauchent entièrement entre :
les petites vésicules exocytaires, qui sont optimisées pour une fusion rapide et temporellement bien contrôlée,
les vésicules plus grandes ou les organites, dans lesquels les barrières énergétiques et les forces d'indentation concomitantes doivent être beaucoup plus grandes.
1. Les courbures membranaires sont différentes.
Les sites de fusion entre deux très grandes vésicules peuvent être considérés approximativement comme plats, ce qui laisse peu de place aux mouvements importants des SNARE et de leurs protéines associées perpendiculairement aux membranes.
Les sites de fusion entre de très petites vésicules, i.e. vésicules synaptiques de diamètre <40 nm, possèdent de très grandes courbures membranaires et offrent beaucoup plus d'espace pour que les complexes SNARE se réorientent eux-mêmes.
Cela peut contribuer de manière significative à réduire la stabilité des pores de fusion, car cela peut faciliter la diffusion des complexes SNARE en dehors du pore de fusion.
2. Une autre différence pertinente concerne la taille de la zone de contact.
a. Cette zone est très grande entre les organites amarrés, i.e. plusieurs μm2 pour les vacuoles de levure.
Le décalage des complexes SNARE volumineux peut imposer la formation de zones de contact plus larges, qui accumulent plus de tension de membrane, qui accélèrent la croissance de la zone de contact et qui permettent à la tension générée de favoriser la fusion avant qu'elle ne puisse se détendre.
L'asymétrie des pores concomitante est due à l'abaissement de l'énergie de flexion de la membrane entre le pore de fusion et le bord incurvé membranaire qui lui est associé, i.e. la présence de complexes volumineux comme HOPS peut augmenter l'énergie libre interface du vertex, en augmentant l'angle de contact entre les vacuoles et en imposant un effet stérique direct sur les membranes.
De plus, la minimisation de la contrainte de courbure grâce à la fusion de petits vésicules peut entraîner l'expansion du pore de fusion, car leur contrainte de courbure peut être importante et compétitive avec la tension de ligne du pore de fusion.
Ainsi, de très petites vésicules devraient être prédisposées pour une progression rapide vers l'ouverture et l'expansion des pores de fusion.






 retrécissement versus effondrement complet).
retrécissement versus effondrement complet).