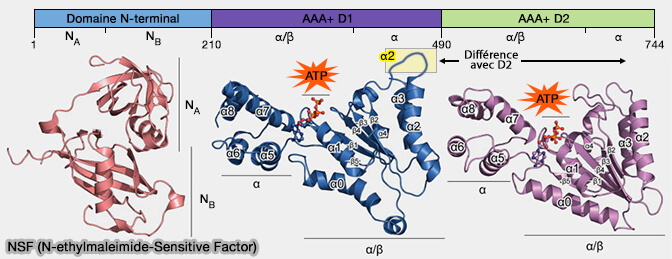
3. Le domaine D2, à activité ATPasique très lente, semble être une protéine d'échafaudage, i.e. principalement utilisée pour la formation et la conservation de la structure hexamérique de NSF.
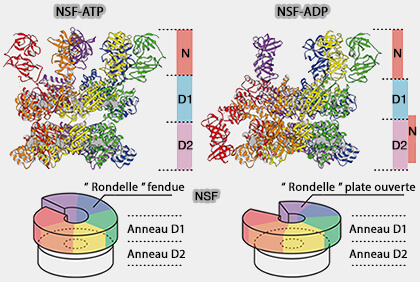
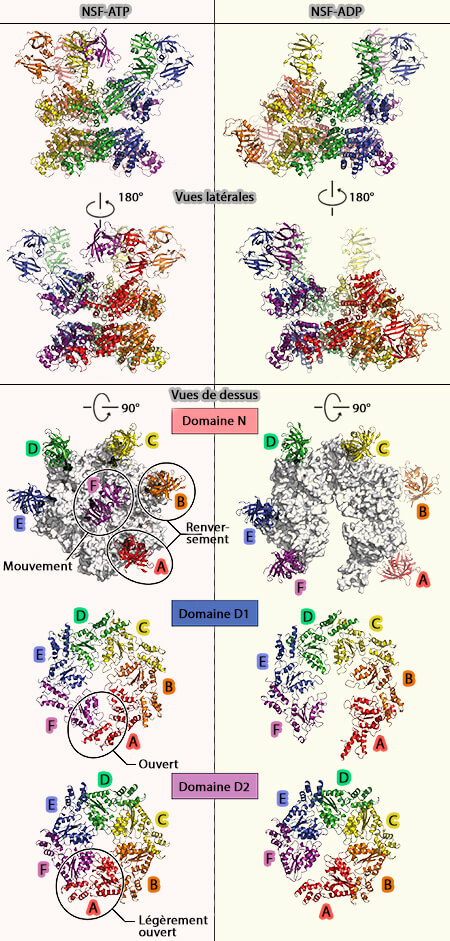
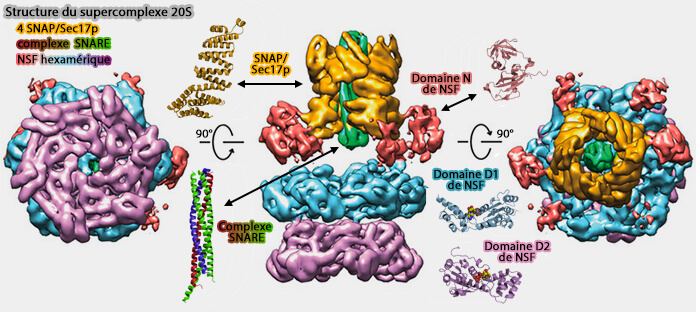
NSF-ATP et NSF-ADP
1. Les structures liées à l'ATP de la NSF sont organisées en trois couches en forme d'anneau des 6 domaines du complexe hexamérique, i.e. 6 N, 6 D1 et 6 D2.
L'anneau D1 possède une conformation en forme de " rondelle " fendue. L'hélice α2 du monomère F est décalée vers le bas par rapport aux hélices α2 des autres domaines.
L'anneau D2 est plat.
2. Les structures liées à l'ADP de la NSF sont organisées aussi en trois couches en forme d'anneau avec une différence de taille, i.e. deux domaines N parmi les six (A et B) semblent être déplacés vers le côté des anneaux ATPases.
L'anneau D1 possède une conformation en forme de " rondelle " plane ouverte, i.e. il est plus étendu et plane et laisse une grande ouverture entre les chaînes A et F.
L'hélice α2 du monomère F est décalée vers le haut par rapport aux hélices α2 des autres domaines (20 Å).
L'anneau D2 laisse un petit espace entre les domaines qui coïncide avec l'espace de l'anneau D1.
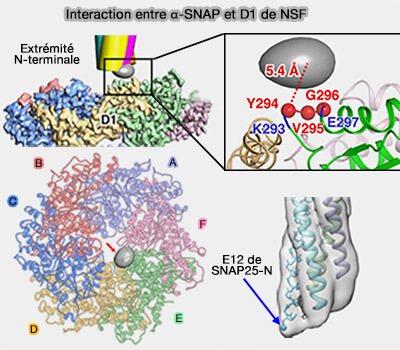
1. Les α-SNAP se lient aux domaines N-terminaux de la NSF, i.e. servant d'adaptateurs entre la NSF et le complexe SNARE, i.e. cette liaison comprime les deux anneaux ATPases de la NSF.
2. La région la plus N-terminale des α-SNAP (E12) se lie au domaine D1, près de la bouche du pore de D1.
Elle semble interagir avec le motif YVG de la chaîne E et chevauche les régions N-terminales du complexe SNARE des six états du 20S.
Interaction entre α-SNAP et D1 de NSF
(Figure : vetopsy.fr d'après Huang et coll)
La distance la plus courte entre les deux est d'environ 5 Å, suggérant fortement une interaction directe entre la région N-terminale du complexe SNARE et la bouche du pore du domaine NSF-D1.
Mécanisme de désassemblage
Plusieurs modèles de mécanismes de désassemblage sont possibles sans forcément s'exclure les uns des autres ou en se complétant, vu les intrications et les interactions des différents membres du complexe 20S.
4. Les interactions renforcées entre α-SNAP et NSF sont plus le fait de liaisons salines que des séquences précises de résidus, ce qui explique qu'un nombre relativement restreint de NSF et de α-SNAP désassemblent un grand nombre de complexes SNARE, i.e plus de 60 chez les mammifères.
Lors de la liaison, les deux anneaux ATPases de la NSF se resserrent comme un ressort comprimé.
Les domaines N sont immobilisés en raison des interactions salines avec les SNAP, ce qui permet de maintenir la tension mécanique.
Changements conformationnels lors de l'hydrolyse de l'ATP
L'hydrolyse de l'ATP apporte alors l'énergie pour initier les changements conformationnels.
Torsion du supercomplexe
1. Les torsions opposées des molécules α-SNAP, i.e. droite, et du faisceau de quatre hélices α du complexe SNARE, i.e. gauche dans les supercomplexes 20S suggèrent qu'une sorte de rotation est impliquée dans le processus de désassemblage du complexe SNARE.
Le supercomplexe 20S exerce un couple de force pour dérouler ou desserrer le complexe SNARE.
2. Une telle rotation a été observée dans l'anneau NSF-D1, qui tourne dans le sens antihoraire par rapport à l'anneau NSF-D2 le long de l'axe central lors de l'hydrolyse de l'ATP.
Dans un complexe 20S, il y a six molécules NSF et quatre molécules α-SNAP, i.e. ce décalage de symétrie implique que les monomères α-SNAP individuels sont liés à un ou deux domaines N-terminaux NSF proches.
Les six états distincts du complexe 20S pourraient présenter un modèle d'interaction unique entre les domaines α-SNAP et les domaines N-terminaux NSF, ainsi qu'une orientation unique du sous-complexe α-SNAP-SNARE par rapport à l'anneau D1D2 de NSF.
3. Une fois liés au substrat, les domaines N de la NSF deviendraient suffisamment stables par rapport à l'anneau D1, permettant ainsi la transmission du mouvement de D1 aux domaines N, et éventuellement au barillet α-SNAP, puisque les domaines N de NSF interagissent directement avec les régions C-terminales des α-SNAP.
Cette rotation du barillet α-SNAP peut générer une force mécanique le long de la direction tangente du canon cylindrique, s'appliquant principalement sur deux sites de VAMP (interactions α-SNAP/SNARE).
Étant donné que l'extrémité N-terminale du complexe SNARE est directement ancrée à la région des pores du D1 de NSF, le couple droit généré lors de l'hydrolyse de l'ATP commencerait à se dérouler et/ou à casser le complexe SNARE hélicoïdal gauche et ainsi, désassembler le complexe en protéines individuelles.
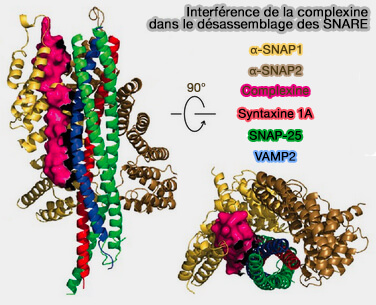
Désassemblage du complexe SNARE
(Figure : vetopsy.fr d'après Huang et coll))
Autres mouvements possibles
1. D'autres forces peuvent agir sur les changements de l'anneau D1.
Un mouvement vertical de 20 Å des boucles de pores peut appliquer une force de cisaillement au complexe SNARE, i.e. l'hélice de torsion droite du domaine D1 de la NSF-ATP, et se détendre pour former une structure plate de la NSF-ADP.
L'ouverture de l'anneau D1, passant d'une conformation en forme de " rondelle " fendue à celle de " rondelle " plate ouverte, appliquerait une force de traction. Les domaines D1 des chaînes A et B tournent vers l'extérieur, de sorte qu'ils pourraient être dans une position privilégiée pour le mouvement des domaines N-terminaux.
2. Le mouvement des deux domaines N (A et B) de NSF, se trouvant au bord inférieur de la rondelle fendue et qui s'inversent dans la NSF liée à l'ADP par rapport à la NSF liée à l'ATP, pourrait appliquer également une force de traction.
Prises ensemble, ces forces désassemblent complètement le complexe SNARE.
Sortie des protéines SNARE
1. L'ouverture de l'anneau D1 dans la NSF liée à l'ADP pourrait servir de sortie pour les protéines SNARE, car un mécanisme de translocation des pores est peu probable ( cf. discussion).
La libération de protéines SNARE et des α-SNAP déclencherait l'échange de nucléotides et redémarrerait le cycle.
2. Le réassemblage du complexe SNARE dans le complexe 20S serait inhibé, évitant ainsi un gaspillage de temps et d'énergie :
par les domaines N-terminaux de plusieurs SNARE,
par séparation physique des protéines SNARE en recyclant les v-SNARE vers la membrane donneuse.
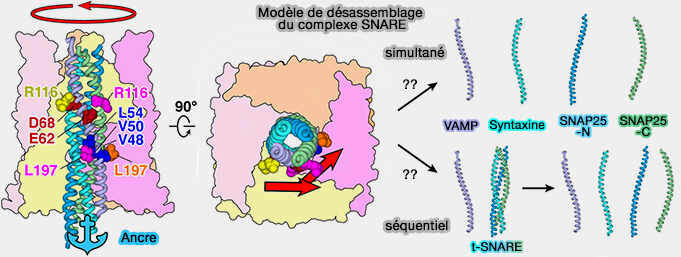
L'une des grandes questions non résolue : les quatre protéines SNARE sont-elles séparées simultanément ou séquentiellement, i.e. sachant que les α-SNAP interagissent principalement sur les VAMP ?
Il semblerait que les α-SNAP aient un rôle supplémentaire dans l'assemblage de complexes SNARE.
Comment et quand les α-SNAP font la différence entre assemblage et désassemblage ?
Comment les α-SNAP font la distinction entre les complexes SNARE normaux ou défectueux ?





 interactions α-SNAP/SNARE).
interactions α-SNAP/SNARE).