1. Le haut degré d'organisation des cellules vivantes qui contraste avec le désordre relatif du milieu extérieur prouve que ces cellules sont capables de transformations accompagnées d'une diminution d'entropie, i.e. mesure du degré de désordre d'un système.
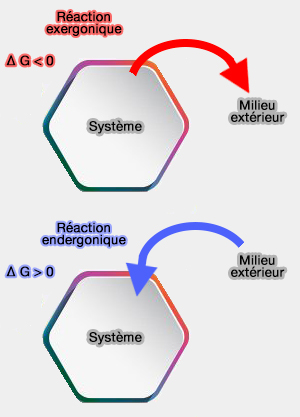
Réactions exergoniques et endergoniques
(Figure : vetopsy.fr)
Ces systèmes doivent donc être associés à d'autres systèmes, dans lesquels se produira, simultanément, un accroissement d'entropie au moins égal en valeur absolue.
Dans les cellules vivantes, on peut considérer que les systèmes chimiques réactionnels se trouvent dans des conditions de réversibilité (température constante) et évoluent à pression et volume constants.
2. La variation d'énergie utilisable sera la variation d'énergie libre ($\Delta G$).
Le système est en équilibre si $\Delta G=0$.
La réaction est dite exergonique si $\Delta G<0$, le système cède du travail au milieu extérieur et de telles réactions peuvent se produire spontanément.
La réaction est dite endergonique si $\Delta G>0$, le système ne peut évoluer que grâce à un apport d'énergie.
3. Dans les systèmes redox, l'énergie apportée par les réactions exergoniques est utilisée par les réactions endergoniques.
Énergie libre standard : énergie de Gibbs
Énergie de Gibbs (G)
1. L'énergie de Gibbs, ou énergie libre de Gibbs (G) est un potentiel thermodynamique utilisé pour calculer la quantité maximale de travail qui peut être extrait d'un système fermé à température et pression fixes, hors le travail dû à la variation de volume.
$G(p, T)=U + pV -TS = H- TS$, équation du système dans laquelle :
$p$ est la pression qui doit être égale à celle de l’environnement pour l’équilibre mécanique.
Vous pouvez lire : Gibbs free energy qui décrit les équations correspondantes.
2. La chaleur de réaction varie avec la température, la pression, la nature et la concentration des corps réagissants.
Pour faciliter les comparaisons, on définit un état standard pour des concentrations molaires, une pression de 1 atmosphère et une température de 25° C.
Par exemple, la phosphorylation du glucose induit une variation d'enthalpie $\Delta H=+ 3 200$ calories, i.e c'est une réaction endothermique.
La combustion du glucose a une $\Delta H=-675$ Kcal, i.e. elle est fortement exothermique.
Variation d'énergie de Gibbs (ΔG)
1. Soit une réaction $\ce{A}$ $\leftrightharpoons$ $\ce{B}$.
a. La constante d'équilibre ($K$) est le rapport des concentrations molaires de B ($[B]$) et de A ($[A]$) à l'équilibre : $K=[B]/[A]$
b. La variation d'énergie libre au cours de la réaction est liée aux concentrations de A et B par la relation de Gibbs-Helmholtz $\Delta G=\Delta G_0+RT\,ln [B]/[A]$ où :
$R$ est la constante des gaz parfaits ( 1.987 cal mole-1 degré-1)
$T$, la température absolue
$\Delta G_0$ est la variation d'énergie libre standard, c'est a-dire celle qui est observée quand A et B sont à une concentration de 1 mole par litre, i.e à ce moment, $\Delta G=\Delta G_0$.
c. Quand la réaction a atteint son équilibre, il n'y a plus de transformation chimique, donc $\Delta G=0$, et à ce moment, le rapport $[B]/[A]$ est égal à la constante d'équilibre $K$, d'où :
a. Pour se rapprocher des conditions biologiques habituelles, on préfère rapporter la valeur de $\Delta G_0$ à des milieux à pH 7, appelée $\Delta G_0'$.
On transforme les ln (népériens) en log (décimaux), i.e. on obtient :
$\Delta G_0'=-1363\,logK$
b. La variation d'énergie libre standard ne dépend que de la constante d'équilibre $K$ du système : sa valeur absolue sera d'autant plus élevée que la réaction sera plus complète dans le sens considéré.
Si la valeur dépasse 1363 x 2 = 2726 en valeur absolue, elle correspond à une valeur de K supérieure à 100 ou inférieure à 1/100, i.e la transformation est pratiquement totale (et irréversible) dans l'un ou l'autre sens.
L'hydrolyse de l'ATP (acide adénosine-triphosphorique) en ADP et $\ce{PO4H3}$ est de - 7 500 calories, cette réaction est exergonique.
Remarque importante : si $\Delta G_0$ est une constante pour une réaction chimique, la valeur de $\Delta G$ dépend des concentrations respectives des composés impliqués dans la réaction.
C'est le signe de $\Delta G$ qui, dans des conditions déterminées, indique si une réaction aura lieu dans un sens ou dans l'autre.
Si les concentrations des réactants et des produits sont telles que $\Delta G$ est négative, la réaction peut se développer même si la valeur de $\Delta G_0$ est positive.
Ceci est très important sur le plan métabolique, car dans les systèmes biochimiques in vivo, les conditions locales sont le plus souvent fort éloignées des conditions standard :
faibles concentrations des composés biochimiques ;
non atteinte de l'équilibre car les produits d'une réaction sont rapidement utilisés ;
systèmes hétérogènes comportant plus d'une phase…
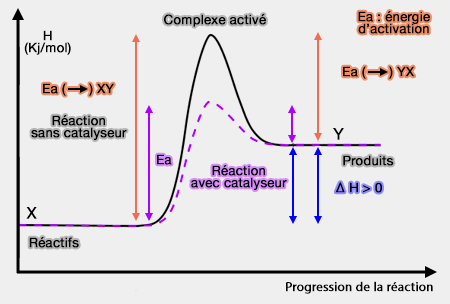
Énergie d'activation : réactions enzymatiques
La condition $\Delta G_0$ n'est pas toujours suffisante pour qu'une réaction ait lieu.
1. On peut apporter au système une certaine quantité d'énergie dite " énergie d'activation ".
Énergie d'activation
(Figure : vetopsy.fr)
Cette énergie peut être fournie en élevant la température par exemple, d'où une méthode de mesure de cette énergie.
2. On peut aussi abaisser le niveau énergétique d'activation, au lieu de fournir de l'énergie au système.
C'est ainsi qu'agissent les catalyseurs ou les enzymes dans les cellules.
Des mécanismes régulateurs permettent de modifier l'activité des enzymes ou leur vitesse de biosynthèse en fonction des besoins cellulaires.
Couplage des réactions
Les variations d'énergie libre sont additives dans une chaîne de réactions consécutives, où chacune utilise le produit de la réaction précédente.
Ceci revient à dire que, dans un système donné, seule compte la variation de $\Delta G$ entre l'état initial et l'état final.
Si une transformation chimique est réalisée en plusieurs étapes, les variations partielles à chaque étape s'ajoutent.
Soit par exemple la suite de réactions :
$\ce{A->B->C->D}$
1. La variation d'énergie libre pour le total des trois réactions, soit $\ce{A->D}$ est la somme algébrique des $\Delta G$ .
Enthalpie et réactions chimiques
(Figure : vetopsy.fr d'après University of Arizona)
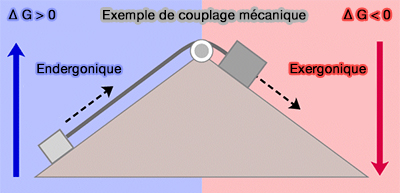
Si cette somme est négative, la transformation de $\ce{A}$ en $\ce{D}$ peut avoir lieu, même si l'une des deux premières réactions a une $\Delta G$ positive, i.e. une réaction fortement exergonique peut " tirer " une réaction endergonique.
Dans une telle succession, les produits $\ce{B}$ et $\ce{C}$ sont appelés " intermédiaires communs " et les réactions sont dites " couplées "
2. Cette notion de réactions couplées présente une importance considérable sur le plan métabolique.
Presque toutes les réactions de l'anabolisme sont endergoniques.
a. Ces réactions à $\Delta G>0$ ne pourra se réaliser que grâce à un apport d'énergie, le plus souvent par intervention d'une réaction exergonique $\Delta G'>0$ et de valeur absolue supérieure à celle de $\Delta G>0$.
b. Par exemple, la transformation du glucose en glucose-6-phosphate, première étape de la glycolyse, est endergonique ($\Delta G>0$) et cette phosphorylation ne peut être réalisée que par couplage avec la rupture d'une liaison pyrophosphate de l'ATP, réaction exergonique, car le bilan final correspond à une $\Delta G>0$.
Bilan : glucose + ATP $\longrightarrow$ glucose-6-phosphate + ADP :
$\ce{\Delta G<0=-4 000}$ cal.mole-1.
Le phosphate n'est pas libéré, mais transféré de l'ATP sur le glucose. De telles réactions dites de transfert seront fréquemment rencontrées dans les métabolismes.






{kind=link}
