La fusion des deux gamètes (spermatozoïde et ovocyte) déclenche l’achèvement de la méiose ovocytaire, la formation transitoire des pronucléi et l’établissement du zygote, première cellule d’un nouvel individu.
La fusion du spermatozoïde avec l'ovocyte provoque une libération massive de Ca++ intracellulaire d'origine ovocytaire (entre 1 à 3 minutes après la fusion) qui provoque :
a. Dans une division classique, une fois que la cellule est arrivée en métaphase, le complexe cycline B/CDK1 doit être inactivé par la dégradation de la cycline B par le complexe APC/C pour le passage en anaphase, puis la fin de la division.
b. Dans l’ovocyte II, cette chute n’a pas lieu spontanément.
La cellule est maintenue artificiellement en métaphase II par un ensemble de mécanismes spécifiques de la méiose ovocytaire, regroupés sous le terme de Cytostatic Factor (CSF).
Ce système est spécifique de la méiose ovocytaire et n’a pas d’équivalent strict en mitose.
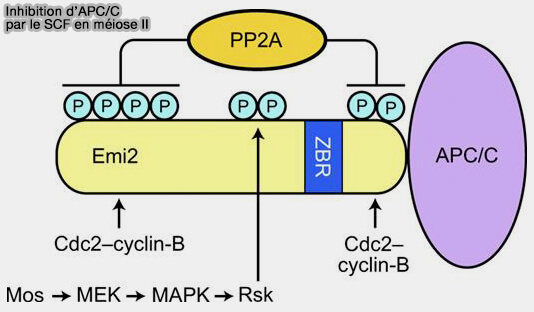
2. Le Cytostatic Factor (CSF) ne correspond pas à une protéine unique, mais à un ensemble coordonné de mécanismes moléculaires qui maintiennent l’ovocyte bloqué en métaphase de la méiose II.
Sa fonction centrale est d’empêcher l’activation effective du complexe APC/C, et donc la dégradation de la cycline B, maintenant ainsi une activité élevée du complexe cycline B/CDK1 (MPF).
a. Emi2 inhibe l’APC/C en se liant directement au coactivateur Cdc20, i.e. mimant les signaux de reconnaissance des substrats de l’APC/C, ce qui permet à Emi2 d’occuper les sites de liaison normalement utilisés par la cycline B et la sécurine.
la kinase Mos, dont le rôle principal est d’activer durablement la voie MAPK dans l’ovocyte,
la voie MAPK (ERK1/2), qui stabilise l’état métaphasique en maintenant l’organisation du fuseau et la stabilité des chromosomes condensés et agissant comme plateforme de signalisation pour activer des effecteurs en aval, notamment p90RSK.
la kinase p90RSK, qui phosphoryle Emi2 sur des sites spécifiques, la protégeant de la dégradation et maintenant sa capacité à inhiber l’APC/C.
Inhibition d’APC/C par le SCF en méiose II
(Figure : vetopsy.fr d'après Wu et Kornbluth)
c. La phosphorylation d’Emi2 par la voie Mos–MAPK–p90RSK stabilise cette interaction, mais le CSF et le MPF ne fonctionnent pas indépendamment et forment une boucle de rétrocontrôle positive :
le CSF empêche la dégradation de la cycline B,
la cycline B maintient CDK1 active,
l’activité CDK1 contribue indirectement à la stabilité des composants du CSF.
d. Dans l’ovocyte bloqué en métaphase II, PP2A agit comme un mécanisme de maintenance en enlèvant continuellement les phosphorylations qui rendraient Emi2 instable, empêchant ainsi l’activation de l’APC/C et la dégradation de la cycline B.
4. Lors de la fécondation, Ca++active principalement la CaMKII (Ca²⁺/calmoduline-dependent protein kinase II),, qui agit en amont en modifiant l’équilibre kinase/phosphatase et en favorisant l’activité de Plx1 sur Emi2, notamment sur des sites chevauchant ou proches de ceux phosphorylés par la voie Mos-MAPK-p90RSK.
Ce processus :
modifie la conformation d’Emi2,
expose ses motifs de reconnaissance par le système de dégradation,
transforme Emi2 d’inhibiteur stable en substrat de l’APC/C.
b. La dégradation d’Emi2 lève progressivement l’inhibition exercée sur l’APC/C, permettant l’ubiquitination et la dégradation protéasomale de la cycline B, la chute de l’activité CDK1 et la sortie de métaphase II.
à l’expulsion d’un deuxième globule polaire identique en taille au premier globule, car le fuseau méiotique est excentré et proche de la membrane plasmique ovocytaire.
Remarque : le site d’expulsion du deuxième globule polaire correspond à une région corticale spécialisée, distincte du site de fusion du spermatozoïde, ce qui limite le risque de perturbation de la ségrégation chromosomique, ce qui évite une aneuploïdie (nombre anormal de chromosomes).
2. Quant au devenir des globules polaires, il dépend du stade méiotique auquel ils sont formés.
Le premier globule polaire, issu de la méiose I, contient un lot haploïde de chromosomes à deux chromatides (1n, 2c), mais ne participe pas à la fécondation et dégénère généralement, bien qu’il puisse, selon les espèces, subir une division équationnelle sans contribution au développement embryonnaire.
Le deuxième globule polaire, formé lors de l’achèvement de la méiose II, contient un lot haploïde de chromosomes à une chromatide (1n, 1c), ne subit aucune division supplémentaire et dégénère rapidement.
3. La maturation ovocytaire est différente chez
la chienne pour plusieurs raisons.
L'ovule ovulé contient encore les 2n chromosomes et n'a pas commencé
la méiose durant la croissance folliculaire, contrairement
aux autres espèces.
L'ovocyte n'expulse ses globules polaires que 48 heures après l'ovulation.
L'ovocyte sécrète de la progestérone avant la lutéinisation (lutéinisation préovulatoire) et, contrairement aux autres
espèces, la reprise de la méiose s'effectue avec des oestrogènes bas et de la progestérone élevée.
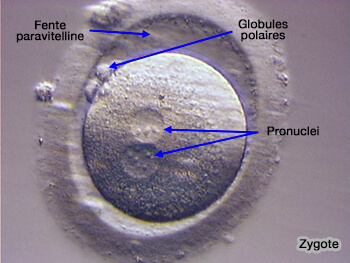
Zygote avec ses deux pronuclei
La maturation ovocytaire s’achève dans la trompe utérine, ce qui suggère un rôle possible des sécrétions tubaires dans la régulation de cette maturation.
Formation des pronuclei
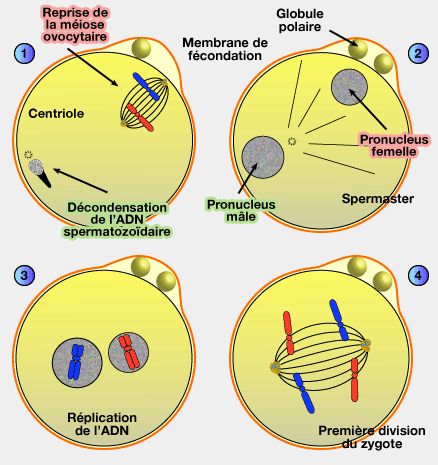
1. Après l’entrée du spermatozoïde dans l’ovocyte II, il n’y a pas de fusion immédiate des noyaux parentaux.
Les deux génomes suivent d’abord des trajectoires distinctes au sein du cytoplasme ovocytaire.
Remarque : nous supposons, sur les schémas, que la cellule somatique ne comporte que 2 chromosomes, donc les gamètes, 1 seul.
a. L’ovocyte II, bloqué en métaphase de la méiose II, possède déjà une organisation nucléaire fonctionnelle.
L’entrée du spermatozoïde déclenche la reprise et l’achèvement de la méiose II, conduisant à la séparation des chromatides sœurs et à l’expulsion d’un deuxième globule polaire.
L’ovocyte devient alors un ovule haploïde, contenant un lot de chromosomes à une chromatide.
Reprise de la méiose II ovocytaire
(Figure : vetopsy.fr)
b. Le spermatozoïde, quant à lui, n’apporte pas une enveloppe nucléaire fonctionnelle.
Son ADN, extrêmement compacté par des protamines, est rapidement décondensé après la fécondation.
L’enveloppe nucléaire d’origine paternelle est désorganisée et éliminée, puis le génome paternel est remodelé par des facteurs d’origine maternelle.
La chromatine spermatique se décondense sous l’action de facteurs d’origine ovocytaire, notamment par réduction des ponts disulfures des protamines. Les protamines sont alors progressivement remplacées par des histones maternelles, selon un processus inverse de celui observé lors de la spermiogenèse.
3. À ce stade, les chromosomes maternels et paternels se décondensent séparément, et deux nouvelles enveloppes nucléaires se forment indépendamment autour de chacun des génomes.
a. Il apparaît alors deux noyaux distincts, le pronucléus femelle et le pronucléus mâle, chacun contenant un génome haploïde et restant physiquement séparé de l’autre.
Le pronucléus mâle se forme progressivement au cours des heures suivant la fécondation (environ 8 à 10 heures chez l’humain) et est généralement plus volumineux que le pronucléus femelle.
Le pronucléus femelle, quant à lui, est localisé plus près des globules polaires, reflet de son origine ovocytaire.
b. La cellule issue de la fécondation constitue déjà le zygote, oeuf fécondé génétiquement diploïde au sens global, mais les deux génomes parentaux restent encore physiquement séparés au sein de pronucléi distincts.
4. Une fois les deux pronucléi établis, chacun réplique son ADN.
Cette réplication correspond à une phase S classique du cycle cellulaire et prépare les chromosomes à la première division cellulaire.
Après cette étape, les deux pronucléi contiennent toujours des génomes séparés, mais désormais sous forme de chromosomes dupliqués.
5. Lorsque la réplication est achevée, les enveloppes nucléaires des deux pronucléi se désassemblent.
Les chromosomes maternels et paternels se retrouvent alors dans un même espace cytoplasmique, sans qu’il y ait eu de fusion nucléaire préalable.
6. Un fuseau mitotique commun se met ensuite en place, sur lequel les chromosomes maternels et paternels s’assemblent et sont répartis ensemble lors de la première division mitotique.
C’est au cours de cette première mitose que chaque cellule fille reçoit, pour la première fois, un noyau contenant à la fois des chromosomes d’origine maternelle et paternelle, établissant ainsi le génome diploïde au niveau nucléaire.
Formation du zygote
Après la disparition des enveloppes pronucléaires, les chromosomes maternels et paternels dupliqués s’assemblent sur un fuseau mitotique commun.
Contrairement à une idée simplifiée, il n’y a pas de fusion directe des noyaux parentaux (caryogamie au sens strict), mais une mise en commun fonctionnelle des chromosomes lors de la première mitose.
1. La première division du zygote correspond à une mitose classique.
Germe et vitellus de
l'oeuf téléolécithe de poule
Elle conduit à la formation de deux blastomères, 24 à 30 heures après la fécondation chez l'homme, à l'origine de l'embryon qui continue sa vie libre dans la trompe utérine.
2. Chez les mammifères placentaires, l'oeuf fécondé est alécithe ou oligolécithe, c’est-à-dire qu’il contient très peu de réserves vitellines,
contrairement aux oeufs télolécithes caractérisés par un volume considérable de vitellus localisé à l'un des pôles,
comme le jaune de l'oeuf de poule.
En raison de cette faible quantité de vitellus, la segmentation du zygote chez les mammifères est holoblastique, c’est-à-dire que la cellule se divise entièrement, sans restriction régionale liée aux réserves. Les divisions successives conduisent ainsi à une augmentation du nombre de blastomères sans augmentation du volume global de l’embryon.
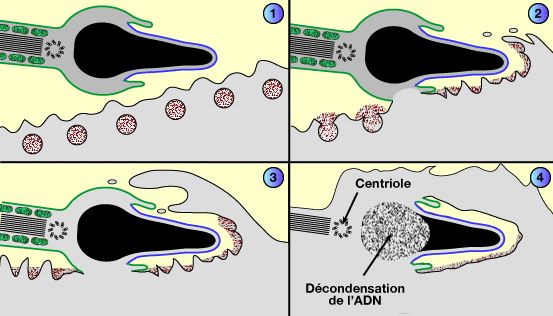
Devenir des organites du spermatozoïde après la fusion
a. Ce centriole forme le spermaster, demi-fuseau méiotique, qui attire le pronucleus femelle vers le pronucleus mâle.
La croissance des microtubules issus du spermaster contribue au rapprochement des deux pronucléi vers le centre de la cellule.
b. Le spermaster disparaît ensuite progressivement.
Lors de la duplication, c'est ce centriole dédoublé qui formera le premier fuseau méiotique du zygote.
3. Tous les autres organites du spermatozoïde, dont les mitochondries sont éliminées par des mécanismes d’autophagie sélective, sont digérés plus ou moins rapidement par le zygote.
L'ADN mitochondrial du nouvel individu est donc un ADN uniquement maternel.