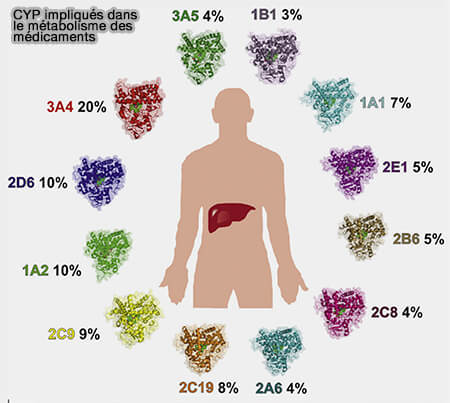
1. Chez l'homme, on trouve 57 gènes CYP fonctionnels. Leurs produits forment 18 familles et 41 sous-familles basées sur la séquence d'acides aminés (Human cytochromes P450 in health and disease 2013).
En 2017, au moins six des 57 CYP humains sont orphelins, i.e. sans substrats identifiables connus.
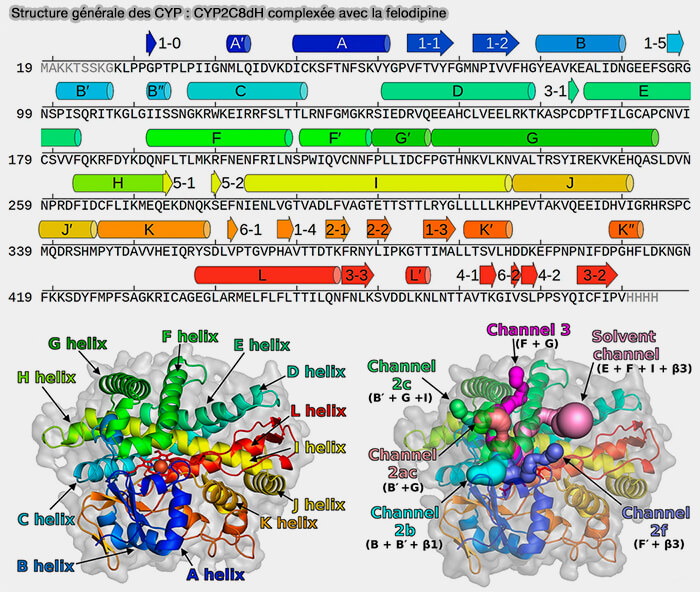
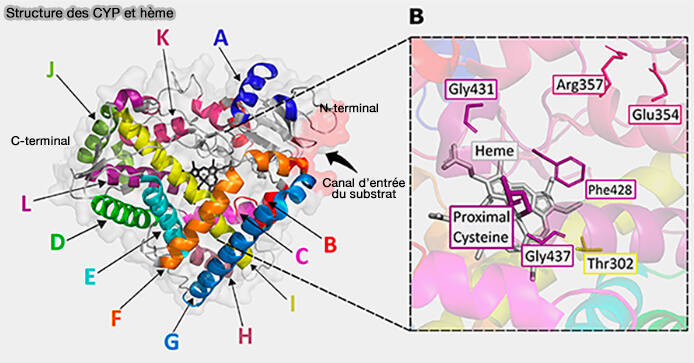
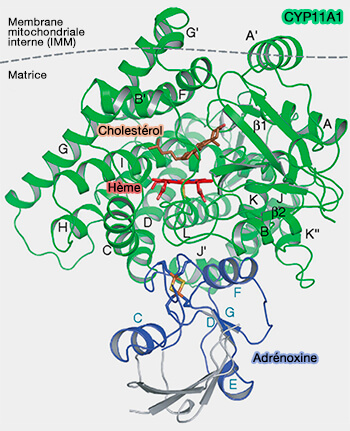
Le domaine catalytique C-terminal, dont la taille et la forme peuvent différer selon les CYP, est principalement composé d'hélices α et abrite l'hème b, i.e. groupe prosthétique, dans une cavité profonde.
Les régions d'hélice B-C et F-G sont les moins conservées et contribuent à la spécificité du substrat, en particulier l'hélice B'.
Ces régions changent de conformation pour l’entrée du ligand et, avec la boucle F-G, forment un toit pour le site actif opposé au plancher de l’hème (What makes a P450 tick 2013).
Structure des CYP : CYP208
(Figure : vetopsy.fr d'après Midlik et coll)
(A/G)XXXT, localisée dans l'hélice I, contient la thréonine hautement conservée qui est un motif de liaison et d'activation de l'oxygène ( réaction 3).
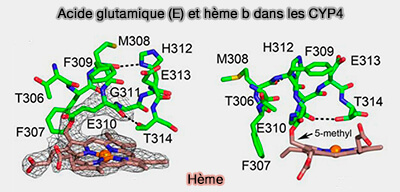
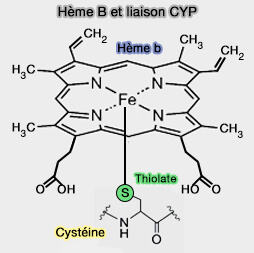
Acide glutamique et hème B dans les CYP4
Figure : vetopsy.fr d'après Hsu et coll)
ExxR contient du glutamate et de l'arginine (R) conservés qui construisent un ensemble d'interactions de ponts salins qui forment la structure tertiaire finale des enzymes.
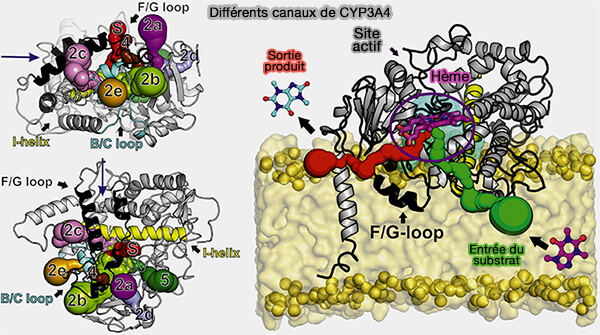
3. Le site actif est profondément enfoui dans la structure et est relié à l'extérieur par un réseau complexe de canaux dans lesquels les substrats et les molécules d'O2 entrent, les produits sortent et les molécules d'eau se déplacent.
b. Les portes d'eau coordonnées contrôlées par les chaînes latérales propionate, i.e. -CH2-CH2-COOH de l'hème et leurs partenaires de pont salin facilitent le mouvement de l'eau.
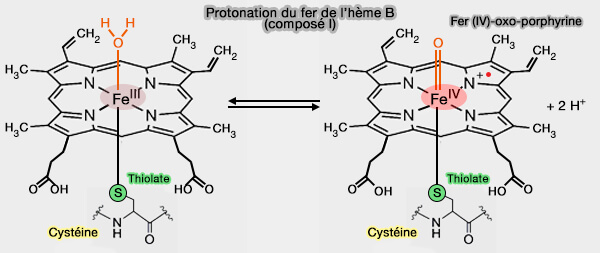
2. L'oxydation des CYP est liée à la formation du composé I, i.e. espèce radicalaire de haute valence, i.e. cation fer (IV)-oxo-porphyrine π ( mécanisme catalytique).
La région de renflement β hautement conservée (FXXGXXXCXG) contenant le ligand thiolate axial de cystéine est rigide de manière à maintenir une distance de liaison hydrogène de deux groupes NH peptidiques (observés dans tous les CYP).
Ces liaisons hydrogène jouent un rôle dans le contrôle du potentiel rédox du fer héminique, qui serait trop faible pour une réduction par les partenaires rédox sans les liaisons hydrogène.
D'autre part, des caractéristiques uniques au sein des sites actifs/canaux d'entrée de différentes enzymes CYP modulent la sélectivité des différents substrats et peuvent entraîner différents produits oxydés du même substrat.
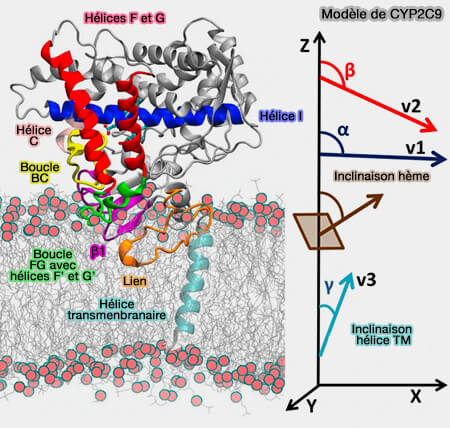
Cette ancre hélicoïdale traverse la bicouche membranaire et est reliée par un lien flexible à un grand domaine globulaire cytosolique qui plonge dans la membrane.
3. L'extrémité N-terminale contient également une séquence peptidique signal pour le trafic vers le compartiment approprié qui, dans le cas des CYP mitochondriaux, est clivée pendant le transport.
Remarque : les CYP des procaryotes ne sont pas des protéines membranaires.




 exemple).
exemple).