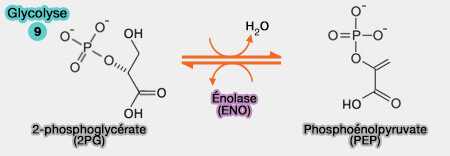
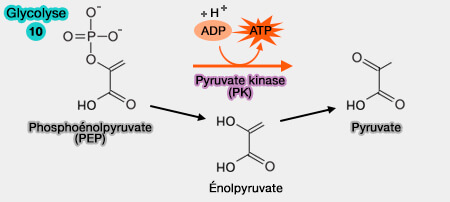
une phase finale, oxydo-réduction couplée à la formation du pyruvate et d'ATP.
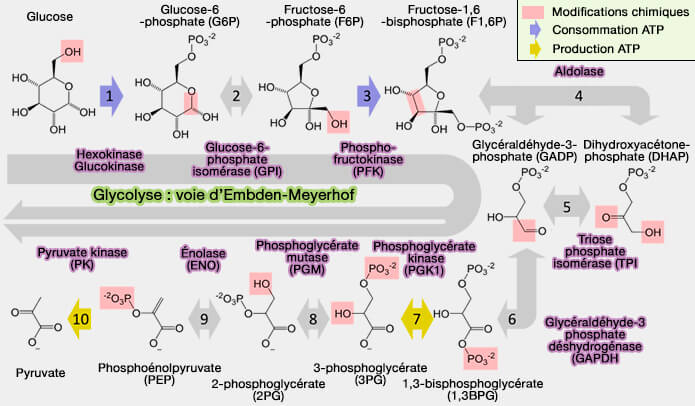
Glycolyse (voie d'Embden-Meyerhof)
(Figure : vetopsy.fr d'après Thomas Shafee)
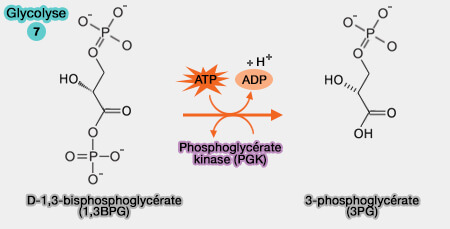
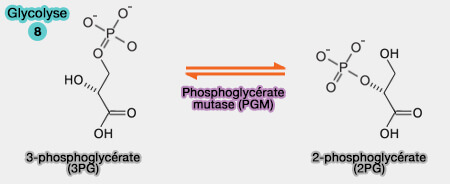
Toutes les réactions sont, en principe, réversibles et équilibrées, exceptées les trois réactions catalysées par des kinases qui sont fortement énergétiques et nécessitent d'autres systèmes enzymatiques pour être inversées :
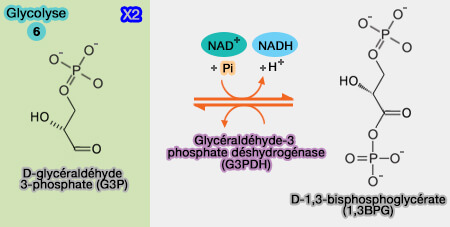
Cette réaction est l'une des étapes les plus importantes de la glycolyse : elle permet la mise en réserve de l'énergie produite par l'oxydation du glycéraldéhyde-3-phosphate, sous forme d'une liaison acylphosphate (composés " riches en énergie ").
Oxydation phosphorylante du glycéraldéhyde-3-phosphate
(Figure : vetopsy.fr)
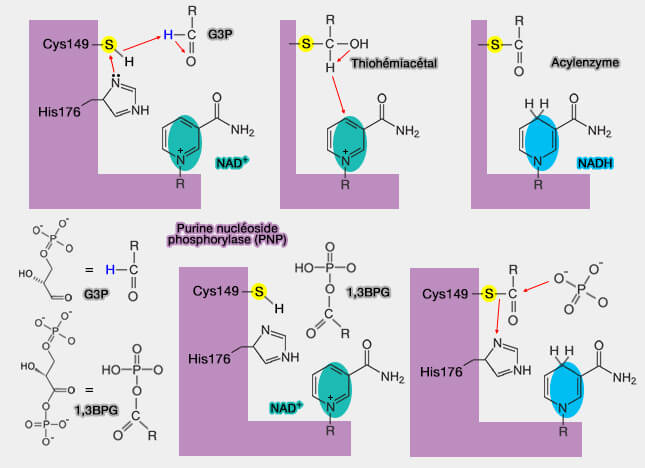
oxydation de ce dernier en thioester ($\ce{R–S–CO–R'$} grâce à l'intervention du NAD+ lié à l'enzyme, ce qui produit un acylthioester qui possède une liaison riche en énergie, i.e. exergonique ($\ce{\Delta G'0}$ d'environ -12 kcal.mole-1), ce qui apporte l'énergie à la réaction suivante ;
Mécanisme de la glycéraldéhyde-3-phosphate déshydrogénase (G3PDH)
(Figure : vetopsy.fr)
Deux isoformes allostériques sont régulées de manière réversible :
PKL, L pour " liver ", foie, mais aussi retrouvé dans les reins et l'intestin grêle,
PKR, R pour " red " globules rouges ou érythrocytes.
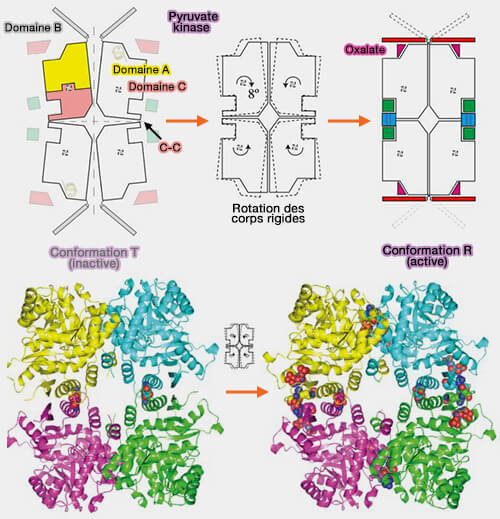
Structure de la pyruvate kinase
(Figure : vetopsy.fr d'après Morgan et coll)
1. La forme active de PKR est un homotétramère dont chaque sous-unité est constituée de quatre domaines :
un petit domaine N-terminal ;
les domaines A et C, impliqués dans les interactions interdomaine ;
le domaine B.
Les sites de liaison du phosphoénolpyruvate et de l'ADP sont situés dans une fente entre les domaines A et B et contiennent les cofacteurs (ions $\ce{Mg++}$ et $\ce{K+}$), essentiels pour la catalyse.
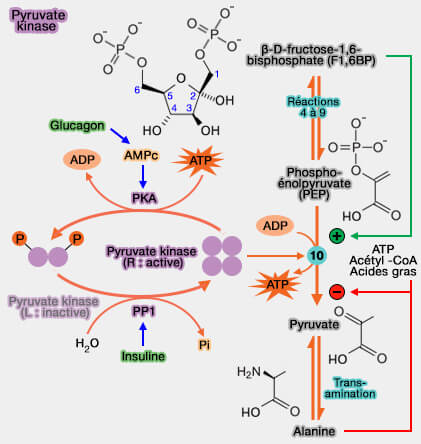
3. Les régulations indirectes sont l'œuvre de réactions de phosphorylation-déphosphorylation de la pyruvate kinase.
Elles économisent la survenue de cycles " futiles " ou cycles du substrat, cycles qui se produisent lorsque deux voies métaboliques se déroulent simultanément dans des directions opposées, dans ce cas glycolyse et gluconéogenèse.
a. Dans le foie, le glucagon et l'épinéphrine activent la protéine kinase A (PKA), qui, à son tour, phosphoryle et désactive la pyruvate kinase, i.e. la glycolyse peut être inhibée en période de pénurie de glucose.
L'activité de la PKA est dépendante des taux cellulaires d'AMP cyclique (AMPc), d'où son autre nom de protéine kinase dépendante de l'AMPc (EC 2.7.11.11).
b. Par contre, la sécrétion d'insuline, suite à l'élévation de la glycémie, active la protéine phosphatase I (PPI), provoquant la déphosphorylation et l'activation de la pyruvate kinase.
c. Le glucagon et l'insuline modulent également la transcription de la PKL.
Isoformes M
Les isoformes dites musculaires (M) sont :
PKM1, retrouvée dans les muscles, mais aussi le cœur et le cerveau, qui n'est pas régulée ;
PKM2, isoforme embryonnaire retrouvée dans les tissus précoces et remplacée progressivement par les autres isoformes.
PKM1 et PKM2 ne diffèrent que de 21 acides aminés, région composée de deux hélices alpha impliquées dans les contacts des sous-unités, déplaçant PKM1 vers une conformation plus active.




{kind=link}
 réaction 1) ;
réaction 1) ;