La régulation du complexe pyruvate déshydrogénase (PDH) doit maintenir l'homéostasie énergétique cellulaire et fournir le carbone nécessaire aux voies de biosynthèse dépendantes du cycle de Krebs.
Lorsque l'acétyl-CoA entre dans le cycle de Krebs, elle produit du NADH et du FADH2 qui sont utilisés pour générer le gradient de protons nécessaire à la phosphorylation oxydative.
La PDH sert de pont entre la glycolyse dans le cytosol et le cycle de Krebs et la phosphorylation oxydative et joue un rôle central dans le métabolisme cellulaire.
La finesse de la régulation de la PDH doit maintenir l'homéostasie énergétique cellulaire et fournir le carbone nécessaire aux voies de biosynthèse dépendantes du cycle de Krebs.
Cette régulation dépend de l'état énergétique de la cellule :
L'ATP, le NADH et l'acétyl-CoA inhibent le complexe, i.e. indicateurs d’un état énergétique élevé de la cellule.
L'ADP, le NAD+, la CoA et le pyruvate l'active, i.e. indicateurs d’un besoin énergétique ou de substrats disponibles.
Régulation rapide du complexe pyruvate déshydrogénase (PDH)
Vue d'ensemble
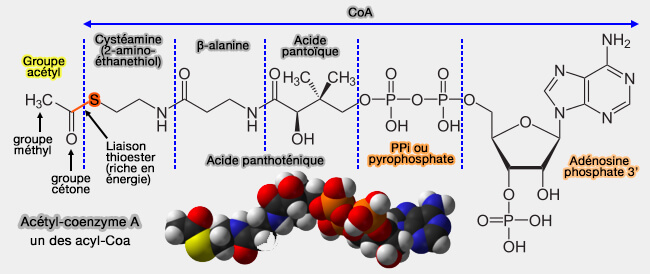
La régulation rapide de l'activité PDH est obtenue par phosphorylation et déphosphorylation de trois résidus sérine, Ser-264 (site 1), Ser-271 (site 2), et Ser-203 (site 3), sur la sous-unité E1α.
Le processus catalysé par E1 est donc l'étape limitante du complexe pyruvate déshydrogénase.
Sites de phosphorylation de E1 par PDK
(Figure : vetopsy.fr d'après Jonathanmott09)
Même si le site 1 (Ser-264) est la cible la plus fréquente, la phosphorylation de n'importe quel site est suffisante pour inhiber l'activité enzymatique.
2. De plus, la PDK elle-même interagit avec le domaine L2, ce qui positionne ce dernier au carrefour entre la régulation, via la phosphorylation, et la catalyse, via le transfert du substrat.
La phosphorylation désorganise donc non seulement l’activité enzymatique d’E1, mais aussi les interfaces fonctionnelles entre E1, L2 et la PDK.
Cette régulation est rapide, réversible, et permet à la cellule d’ajuster le métabolisme énergétique en fonction des besoins.
1. Quatre isoformes de PDK ont été trouvées (PDK1, PDK2, PDK3 et PDK4) qui varient en taille, dans leur localisation cellulaire, mais qui se chevauchent souvent.
On note des différences entre les isoformes dans les cinétiques de réaction, les sites de phosphorylation de E1, ses affinités de liaison pour les domaines lypoyles.
En effet, l'interaction de la PDK requiert la présence du domaine lypoyle L2 sur E2 pour stabiliser l'interaction PDK–E1 et favoriser une conformation qui rend les sites de phosphorylation accessibles.
Le domaine L1, plus distal, semble jouer un rôle secondaire dans cette interaction.
Reins, cerveau, foie, présente aussi dans le muscle
Induite en condition
de jeûne, diabète, exercice
Régulée par des facteurs hormonaux : glucocorticoïdes, insuline (répression), PPARs (activation)
Favorise la gluconéogenèse et la conservation du glucose
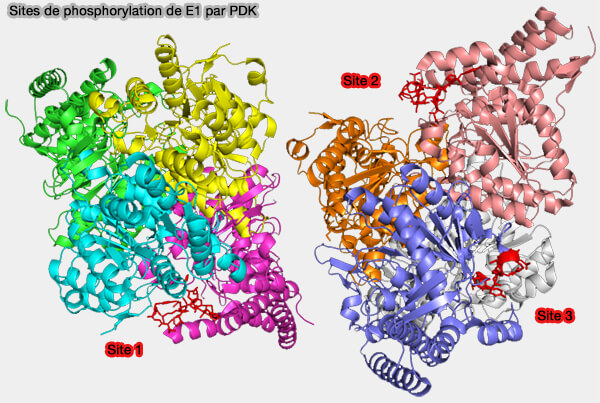
a. Chaque sous-unité de la PDK comprend deux domaines structuraux séparés par une fente.
Structure du complexe PDHK2-L2
(Figure : vetopsy.fr d'après Devedjiev et coll)
Le domaine N-terminal régulateur, i.e. domaine R, est responsable de l'interaction avec le domaine lipoyle L2 du complexe PDH,
Le domaine C-terminal catalytique, i.e. domaine K, est impliqué à la fois dans l’activité kinase et dans la dimérisation de l'enzyme.
La fente située entre ces deux domaines forme le site actif, dans lequel s’insèrent l’ATP et le segment de la sous-unité E1 contenant la sérine cible.
a. PDK doit donc se déplacer sur toute la surface de PDH pour l'inactiver complètement.
Lorsque l'une des sous-unités PDK est liée à L2 (interne), l'auttre sous-unité serait libre de se mouvoir.
Cette organisation asymétrique permettrait à la PDK d'osciller ou de pivoter sur le complexe PDH, en utilisant les différents domaines L2 comme points d’ancrage successifs.
Ce mécanisme souligne l’importance structurale des domaines L2, non seulement comme partenaires transitoires, mais aussi comme guides dynamiques pour la régulation du complexe.
3. Les régulations de la LDH sont dues aux régulations de la PDK : comme la PDK inhibe la PDH, elles sont strictement inverses (cf. plus haut).
Pyruvate déshydrogénase phosphatase (PDP)
La pyruvate déshydrogénase phosphatase (EC 3.1.3.43) catalyse la réaction inverse de celle de la PDK en éliminant la phosphorylation des sérines de la sous-unité E1 du complexe PDH, ce qui entraîne la réactivation du complexe.
Elle augmente le flux d'acétyl-CoA dans le cycle de Krebs pour favoriser la phosphorylation oxydative et la biosynthèse.
Structure
1. Deux isoformes de PDP ont été trouvés chez l'homme.
PDP1 est fortement exprimée dans le cerveau, le cœur, le muscle squelettique et les testicules, régulant le métabolisme énergétique dans les tissus à forte demande d’énergie.
PDP2 est fortement exprimée dans le foie et le tissu adipeux, contrôlant le métabolisme énergétique dans les tissus de stockage et de détoxification.
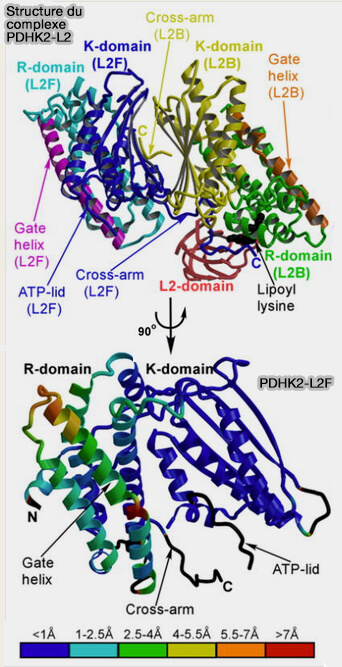
a. La sous-unité catalytique (PDP1c) appartient à la famille des phosphatases sérine/thréonine PP2C (phosphatase 2C).
Une poche hydrophobe spécifique à la sous-unité catalytique de PDP1, i.e. résidus 98-51, est probablement importante pour la liaison d’un ligand, d’un domaine lipoylé ou d’un autre facteur.
Le " flap segment ", segment en forme de languette, est une partie flexible de la protéine qui peut se déplacer pour ouvrir ou fermer cette poche, ou pour permettre l’interaction avec des substrats ou des partenaires.
L’activité de PDP1 est stimulée en réponse à une élévation du calcium dans les cellules musculaires, mécanisme essentiel lors de la contraction musculaire.
Le PDH ainsi activé renforce la production de NADH et d’ATP mitochondriale en réponse directe à la demande énergétique accrue du muscle en contraction.
Remarque : L'activité PDP2, non activée par le calcium, est augmentée en présence de spermine, sans que l'on comprenne sa signification.
2. La régulation post-traductionnelle impliquerait aussi une phosphorylation par la protéine kinase C delta (PKCδ), ce qui entraîne une augmentation de leur activité phosphatase, i.e. renforcement de l’activité du complexe PDH.
1. Cette altération entraîne une accumulation de pyruvate, qui est alors redirigé vers la voie anaérobie, menant à une accumulation de lactate, ce qui provoque :





{kind=link}

 charge énergétique de la cellule).
charge énergétique de la cellule).