Bassoon et Piccolo, deux très grandes protéines d’échafaudage de la cytomatrice de la zone active (CAZ), ont de nombreux rôles, dont celui de Piccolo sur le cytosquelette d'actine.
Molécules de la CAZ et cycle vésiculaire
(Figure : vetopsy.fr d'après Gundelfinger et coll)
Suite des fonctions de Bassoon et Piccolo
Rôle spécifique de Piccolo et dynamique de l'actine
Alors que Piccolo et Bassoon partagent une grande quantité d'homologie structurelle, chacun contient également des segments uniques, ce qui implique des fonctions divergentes.
Dans le recrutement dans la ZA
Piccolo est sélectivement nécessaire à :
l'assemblage des réseaux d'actine induits par l'activité au sein de boutons présynaptiques,
Fonctionnellement, ces interactions semblent contribuer à la régulation de la translocation et du recyclage des vésicules synaptiques (VS) au sein des terminaux nerveux.
Structures et interactions de Bassoon et Piccolo
(Figure : vetopsy.fr d'après Gundelfinger et coll)
Interactions Piccolo/Daam1
(Figure : vetopsy.fr d'après Wagh et coll)
3. Piccolo semble agir comme une plate-forme pour la polymérisation de ces filaments.
a. Daam1 se lie à une région située entre CC1 et CC2 de Piccolo, i.e. Pclo1980-2553.
Cette région n'est pas conservée chez Bassoon, mais interagit avec d'autres protéines associées à l'actine, notamment GIT1 et la profiline1/2, i.e. cette région de Piccolo a un rôle unique dans l'assemblage de l'actine-F présynaptique.
Remarque : Daam1 est normalement inactivé par l'interaction entre son domaine GBD et son domaine DAD, qui courbe la protéine et enterre les domaines FH1 et FH2.
Un exemple d'activation de Daam
(Figure : vetopsy.fr d'après Liu et coll)
Ce n'est qu'après son activation que Daam1 se lie à Piccolo et favorise l'assemblage de l'actine-F, ce qui implique que Piccolo contrôle spatialement l'assemblage de ces filaments à partir de la ZA.
GIT1, et éventuellement Piccolo chez les vertébrés, aident à coupler l'exocytose et l'endocytose en créant un pont entre les zones active et endocytaire.
Ce processus pourrait être facilité par l'intermédiaire de complexes entre Piccolo, GIT1, Abp1 et d'autres protéines d'assemblage de l'actine au niveau des sites endocytaires.
Entretien et intégrité des synapses
En plus de leur rôle dans l'assemblage structurel et fonctionnel de la zone active (ZA) présynaptique, Piccolo et Bassoon sont impliqués dans :
une réorganisation moléculaire de la machinerie de libération, i.e. (1) synthèse des protéines et (2) dégradation des protéines via le système ubiquitine-protéasome (UPS).
b. Le silence présynaptique induit par la dépolarisation est caractérisé par :
une diminution de la taille du RRP et une diminution de la pr, i.e. probabilité de fusion d'une vésicule synaptique (VS) amarrée,
L'induction/récupération du silence présynaptique implique également la régulation de l'activité de l'adénylate cyclase (AC), qui est un régulateur en amont de la PKA.
La liaison Siah1 aux domaines de Bassoon et de Piccolo inhibe son activité.
c. Ce système de dégradation important pourrait être impliqué dans :
la libération régulée de neurotransmetteurs,
le recyclage des VS,
et/ou l'élimination des protéines mal repliées ou vieillissantes.
Voie de signalisation présynapse/noyau
La communication entre les synapses et le soma, en particulier le noyau, d'un neurone est essentielle pour la survie neuronale ainsi que pour les processus de plasticité homéostatique et associative.
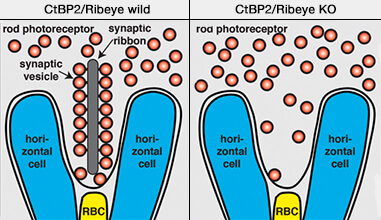
Délétion de Ribeye dans les synapses à ruban
(Figure : vetopsy.fr d'après Maxeiner et coll)
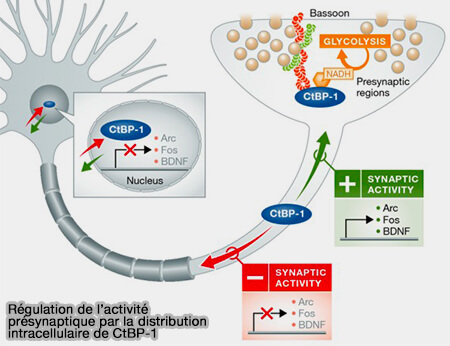
L'interaction entre Bassoon ou Piccolo et CtBP1 est contrôlée par NAD/NADH et contribue à la distribution de CtBP1 dépendante de l'activité entre les présynapses et le noyau.
L'activité élevée fait évoluer l'équilibre entre les pools synaptiques et nucléaires CtBP1 en synapses, faible activité vers le noyau (Presynapses go nuclear! 2015).
Bassoon et Piccolo contribuent de manière significative à l'ajustement de l'expression génique régulée par l'activité et, à leur tour, à la mise en œuvre à long terme des mémoires.
3. Les CtBP travaillent avec différentes protéines d'échafaudage comme Bassoon et Piccolo pour contrôler la fission de la membrane vésiculaire, l'assemblage de rubans synaptiques et l'expression des gènes nucléaires ( rôles de ribeye dans les synapses à ruban).
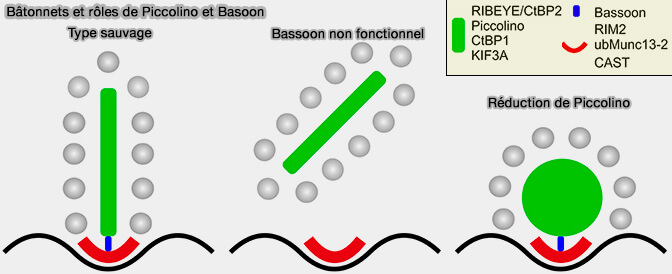
Bâtonnets et rôles de Piccolino et Basoon
(Figure : vetopsy.fr d'après Regus-Leidig et coll)



{kind=link}


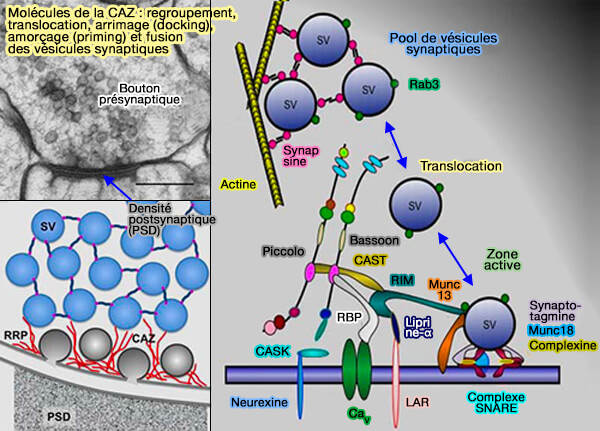

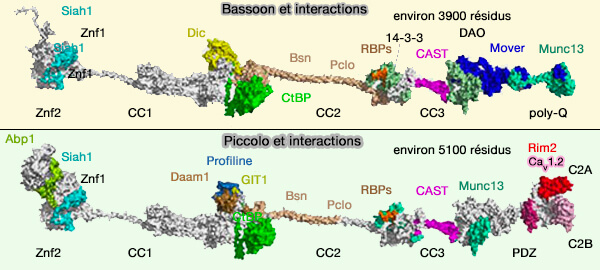
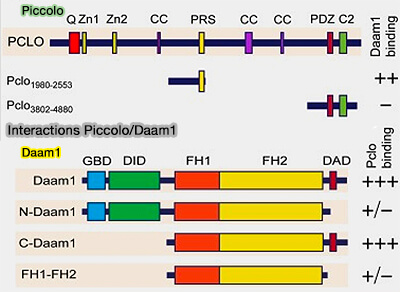
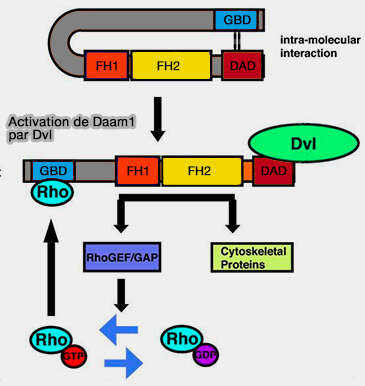
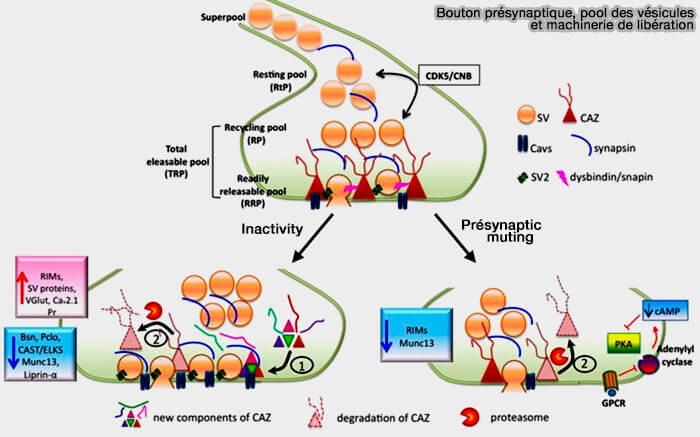
 tableau des demi-vies des protéines synaptiques).
tableau des demi-vies des protéines synaptiques).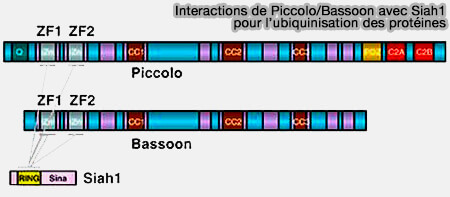

 rôles de ribeye dans les synapses à ruban).
rôles de ribeye dans les synapses à ruban).