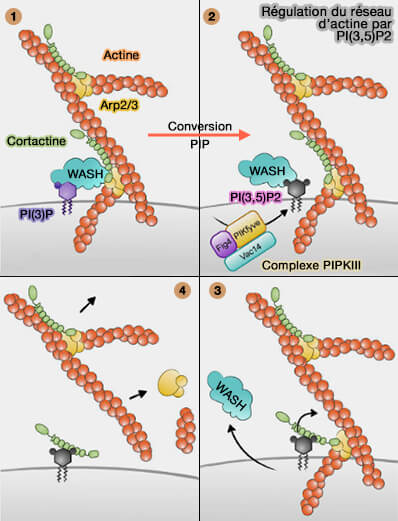
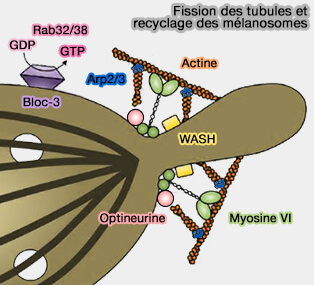
La fission lysosomale peut provoquer la formation :
Fission des lysosomes
(Figure : vetopsy.fr d'après Trivedi et coll et Saffti et coll)d'une vésicule détachée directement à partir du lysosome ou à partir de l'extrémité d'un tubule du lysosome,
2. Les lysosomes peuvent entrer de manière transitoire en contact par leur membrane plasmique avec d'autres organites, i.e. par un petit pore de fusion qui permet l'échange de molécules (kiss), et se séparent (run) rapidement (modèle " kiss-and-run "), i.e. ce processus nécessite une fusion temporaire et une fission.
La clathrine provoquerait la courbure initiale de la membrane pour former un bourgeonnement lysosomal, bourgeonnement qui est ensuite allongé pour former un tubule plutôt qu'une vésicule (Kinesin 1 Drives Autolysosome Tubulation 2016).
Vue d'ensemble
1. Les lysosomes produisent des tubules qui peuvent être sectionnés soit au niveau de leur cou, soit à leur extrémité distale.
2. La formation des tubules nécessite trois étapes.
a. La déformation membranaire nécessite une protéine de manteau, par exemple la clathrine, pour induire la courbure de la membrane et la naissance de l'extrémité du tubule naissant.
b. La pointe du tubule s'allonge par extrusion membranaire du lysosome parent, nécessitant probablement des protéines motrices générant des forces et/ou des protéines qui maintiennent la courbure comme les nexines.
c. Les tubules allongés subissent une fission pour former des transporteurs intermédiaires, nécessitant des machineries de scission et/ou de constriction comme les complexes de dynamine et d'actomyosine ( actomyosine et fission des lysosomes).
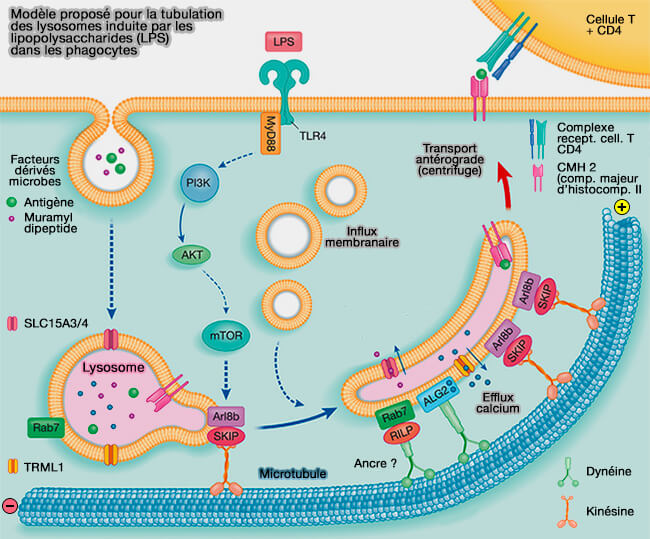
Modèle pour la tubulation des lysosomes induite par les LPS dans les phagocytes
(Figure : vetopsy.fr d'après Hipolito et Coll)
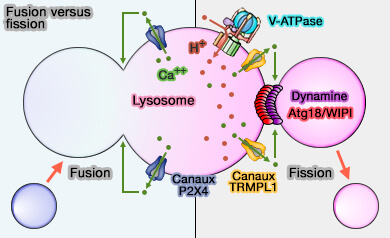
Fusion versus fission des lysosomes
(Figure : vetopsy.fr d'après Saffti et coll)
Son activation réduit la taille des lysosomes dans les cellules dans lesquelles PIKfyve a été inhibé.
Le Ca++ libéré pourrait stimuler la calmoduline (CaM) et l'activité de la dynamine-1, bien que les neurones Fab1 -/- présentent un taux réduit de dynamine-1 pour des raisons inconnues.
Ce double rôle du Ca++, fission et fusion, pourrait être subordonné aux gradients de pH des microdomaines de la membrane lysosomale.
Une forte concentration en H+ favoriserait la fission via TRPML1.
Une faible concentration en H+, par réduction de l'activité de la V-ATPase, favoriserait la fusion via P2X4.
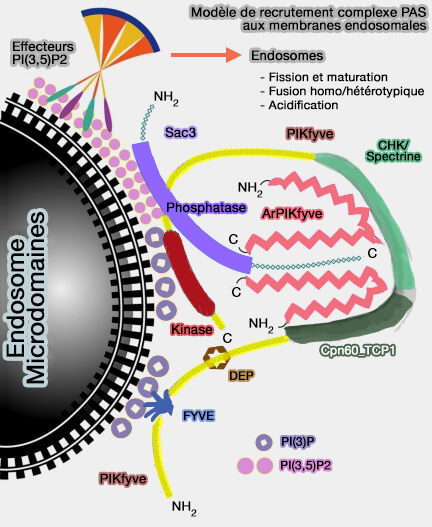
b. PI(3,5)P2 pourrait également réguler la fission de lysosomes par la modulation des protéines de la famille des PROPPIN, i.e. ATG18, ATG21 et HSV2 dans la levure et WIPI-1 à WIPI-4 chez les mammifères.
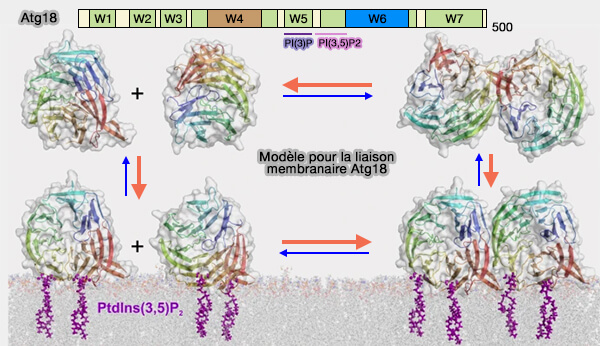
ATG18 pourrait provoquer la fission en insérant une boucle désorganisée dans les membranes pour former une α-hélice amphipathique qui amène ATG18 près de PI(3,5)P2 pour favoriser l'oligomérisation d'ATG18 et la déformation de la membrane (Membrane scission driven by the PROPPIN Atg18 2017).
ATG18 pourrait alors fonctionner avec des protéines dynamine-like comme Vps1 pour induire la fission.
Atg18 et sites de liaison membranaire
(Figure : vetopsy.fr d'après Scacioc et coll)
Le Ca++ libéré par TRPML1 et PI(3,5)P2 pourrait ainsi réguler les PROPPIN pour stimuler la fission.
b. On ne sait pas si des processus de fission similaires s'appliquent aux lysosomes, bien que, dans les marqueurs endosomaux utilisés, on trouve Rab7 associée aussi bien aux endosomes tardifs qu'aux lysosomes.




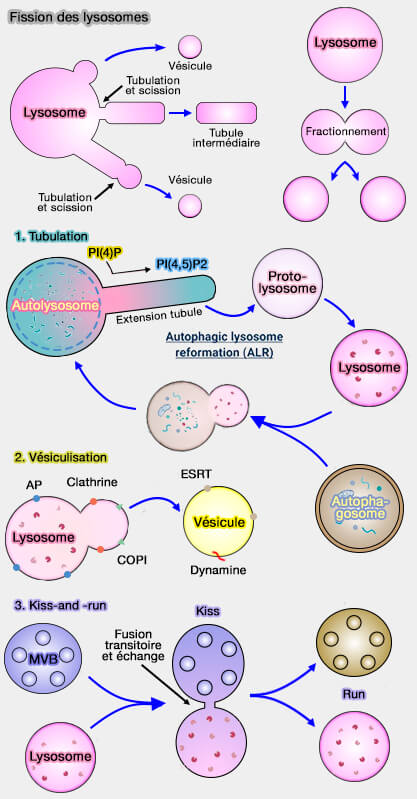
 modèle " kiss-and-run "), i.e. ce processus nécessite une fusion temporaire et une fission.
modèle " kiss-and-run "), i.e. ce processus nécessite une fusion temporaire et une fission.