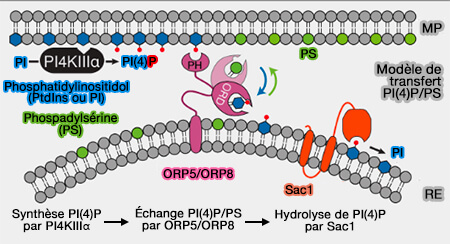
Modèle de transfert de PI(4)P/PS par ORP5/8
(Figure : vetopsy.fr d'après Chung et coll)
Comme ces ORP sont ancrés dans le RE, ils n'ont pas besoin de se lier au VAP pour réguler le trafic des lipides.
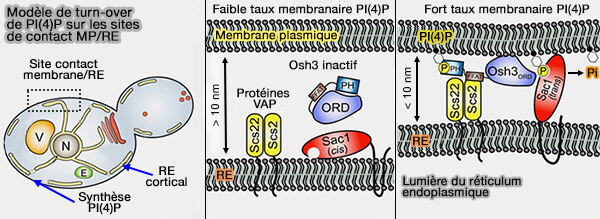
2. Osh3 de la levure régule le métabolisme de PI(4)P au niveau des MCS RE/PM, mais a besoin d'un domaine FFAT pour se lier aux orthologues de VAP, Scs2/Scs22 pour l'amener au RE.
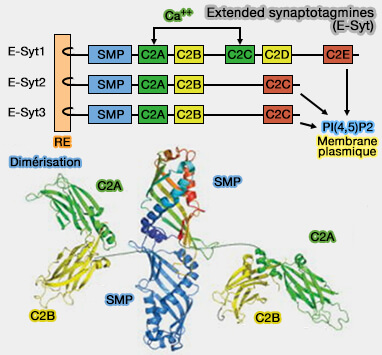
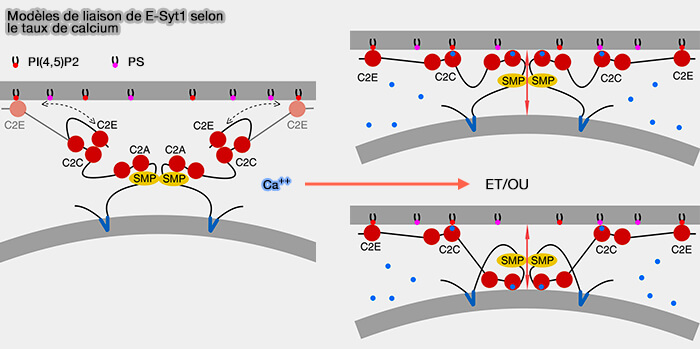
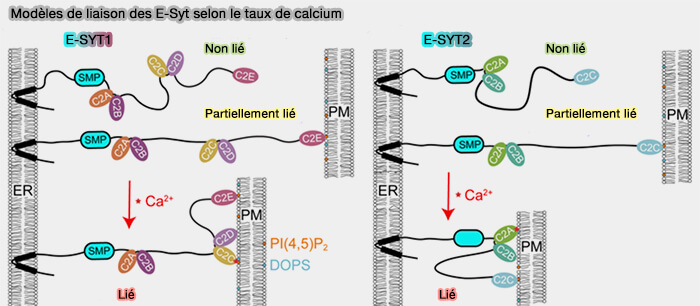
Certains suggèrent que les domaines C2 s'attachent tous sur la membrane plasmique.
D'autres que les premiers domaines sont liés à la membrane du réticulum endoplasmique.
Transfert de lipides et modèles
1. Les domaines SMP dimérisés transportent les glycérophospholipides entre le RE et la MP, selon leurs gradients de concentration, en utilisant un modèle d'extraction-glissement-diffusion-libération.
Les longueurs des liens et les dimensions des protéines fournissent la liberté conformationnelle nécessaire.
Le transfert par pontage direct des deux organites par le dimère des domaines tubulaires SMP est totalement exclu.
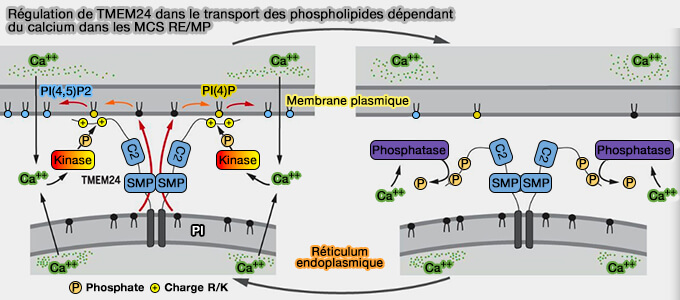
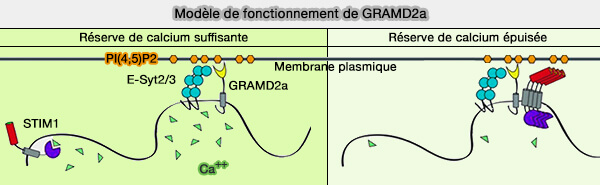
TMEM24 est une protéine du RE qui régule le transfert du phosphatidylinositol (PtdIns ou PI) au niveau des MCS RE/PM en fonction des changements de concentration cytosolique de Ca++.
un domaine C-terminal qui se lie à la membrane plasmique.
Régulation de TMEM24
(Figure : vetopsy.fr
d'après Lees et coll)
2. TMEM24 transfère préférentiellement le phosphatidylinositol du réticulum endoplasmique (RE) à la membrane plasmique (PM), i.e. ce qui provoque la diminution de la sécrétion d'insuline lors de la stimulation du glucose.
La régulation de TMEM24 dans les MCS RE/PM dépend de la concentration de Ca++.
Lorsque la concentration calcique cytosolique augmente, TMEM24 est phosphorylée par PKC et sa concentration diminue dans les MCS RE/MP.
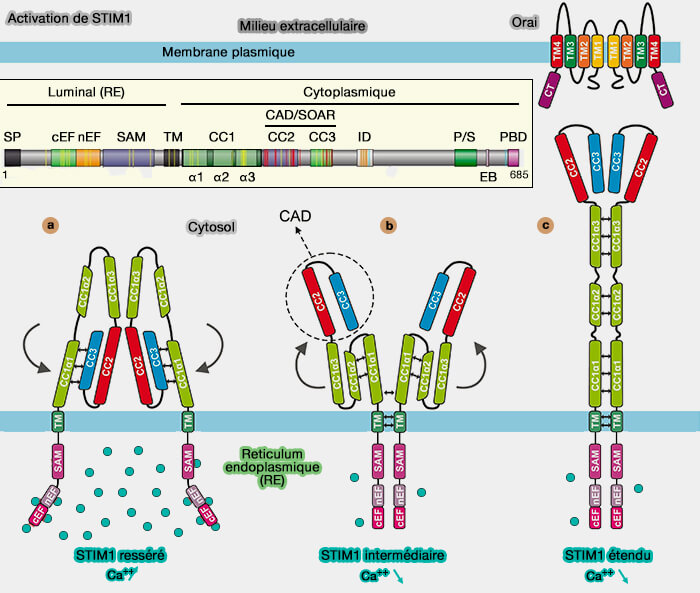
2. STIM1 (STromal Interaction Molecule1) est une protéine membranaire dimérique du RE qui comprend :
un seul domaine TM,
un domaine cytosolique qui comprend une région polybasique responsable de la liaison avec les lipides de la MP,
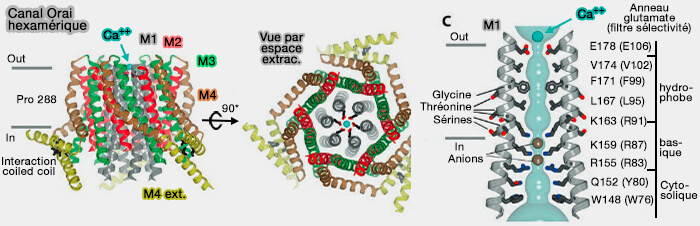
trois domaines coiled coil, i.e. CC1, et CC2/3 appelés aussi CAD ou Crac Activation Domain ou SOAR ou STIM-Orai Activating Region nécessaires à l'activation d'Orai1, (Mechanism of STIM activation 2020)
a. À l'état de repos, lorsque la réserve de calcium du réticulum endoplasmique (RE) est pleine, un sous-domaine de CC1 (appelé α1) et les domaines CC2-CC3 de CAD forment une interaction coiled coil en spirale (représentée par de petites flèches doubles noires), ce qui empêche la liaison à Orai (Cell Signaling: The many states of STIM1 2021)
b. L'épuisement des réserves du Ca++ du RE peut être dû à des facteurs de croissance, des hormones ou des molécules étrangères qui peuvent déclencher des voies de signalisation dépendantes du calcium.
Dès lors, STIM1 à l'intérieur du RE (rose) perd ses ions Ca++ liés (sphères vertes), provoquant l'interaction des régions entre elles et provoquant une interaction (doubles flèches) entre les domaines transmembranaires (TM vert foncé) des protéines STIM dimérisées.
Ces réarrangements conduisent probablement à un état intermédiaire dans lequel l'interaction coiled coil CC1α1-CAD est libérée (représentée par des flèches courbes), et la CAD est projetée vers la membrane plasmique tandis que CC1α1 se rapproche des deux autres sous-domaines de CC1 (α2 et α3).
c. STIM1 allongée s'étend dans les MCS RE/MP pour rejoindre la MP où elle se lie et active Orai1 et déclenche l'ouverture du canal et permettant aux ions calcium d'entrer dans la cellule (Store-Operated Calcium Channels 2015).
Remarque : dans les cellules musculaires squelettiques, les MCS sont composées du récepteur Ryanodine1 (RyR1) sur la citerne terminale du RE et Cav1.1 sur la membrane plasmique, RyR2 et Cav1.2 dans le muscle cardiaque.
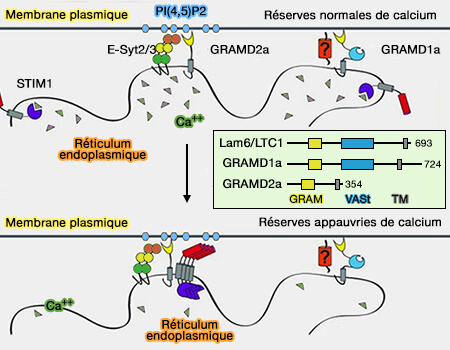
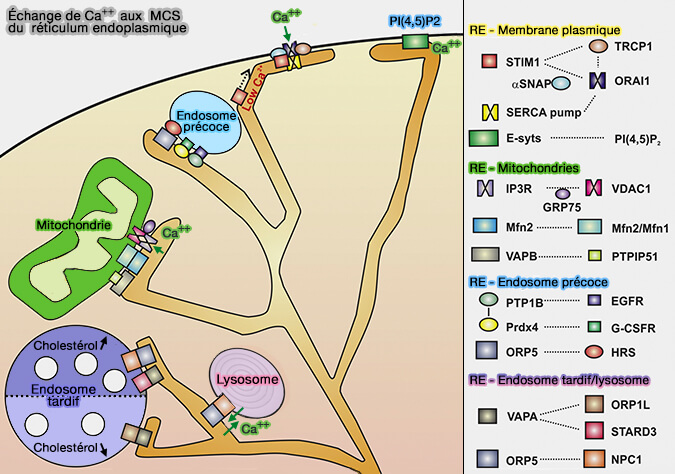
Échange de Ca++ aux MCS du réticulum endoplasmique
(Figure : vetopsy.fr d'après Burgoyne et coll)





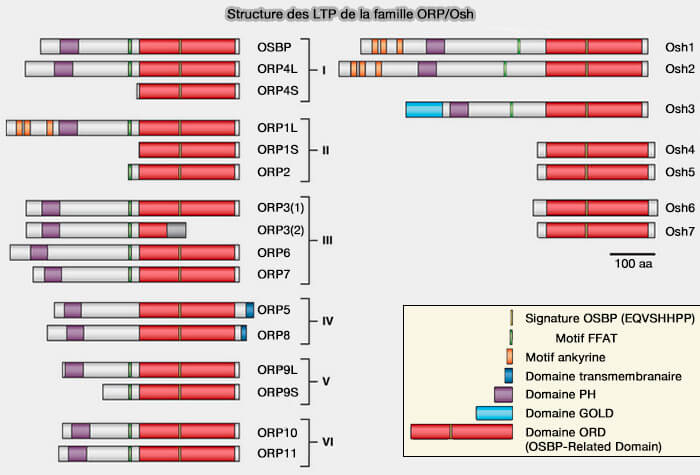
 structure de OSBP).
structure de OSBP).