Bioénergétique : composés " riches en énergie "
Composés à liaison phosphate
ATP : phosphorylation oxydative
ATP synthase : vue d'ensemble du mécanisme
L'ATP synthase est un moteur rotatif biologique qui utilise un mécanisme catalytique rotatif pour coupler la translocation des protons à travers une membrane avec la synthèse de l'adénosine triphosphate (ATP) à partir du phosphate inorganique (PI) et de l'adénosine diphosphate (ADP).
Chez les eucaryotes, ce processus est localisé en totalité dans les mitochondries, au niveau de la membrane interne (IMM), ou dans les chloroplastes, mais dans la membrane plasmique chez les bactéries.
Vue d'ensemble du mécanisme
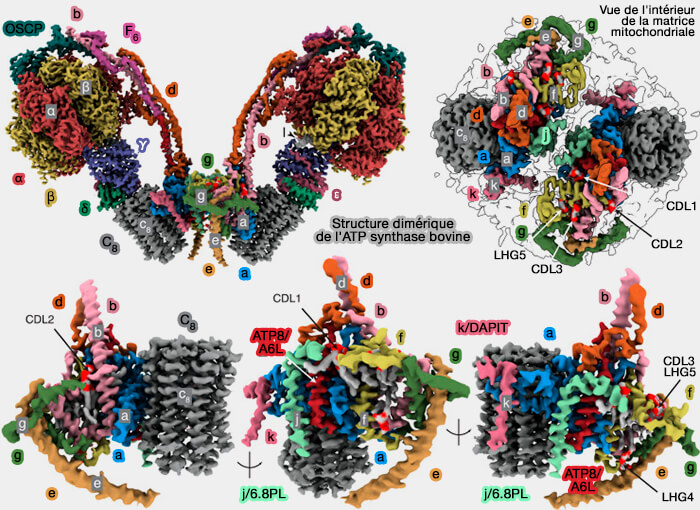
Structure dimérique de l'ATP synthase bovine
(Figure : vetopsy.fr d'après Spikes et coll)
Vue d'ensemble du mécanisme
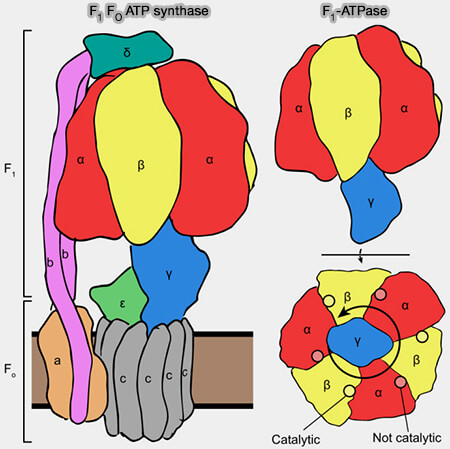
L'ATPase synthase comprend deux moteurs moléculaires, F1 et FO, couplés ensemble par une tige centrale et une tige périphérique.
1. L’ATP synthase est constituée de deux régions principales qui sont des moteurs rotatifs.
Ce rotor central entraîne des changements conformationnels dans la F1-ATPase catalytique, où l'ATP est synthétisé à partir de l'ADP et de PI.
2. F1 et FO sont couplés par :
une tige centrale qui tourne et qui transmet l'énergie de rotation du moteur de FO au domaine F1,
une tige périphérique (PS), i.e. sous-unités OSCP, b, d, F6 et la région extramembranaire d'ATP8/A6L statique qui relie la surface externe du domaine catalytique à la sous-unité a dans le domaine membranaire.
Le domaine (αβ)3, la tige périphérique (PS) et la sous-unité a/ATP6 constituent le stator de l’enzyme contre lequel tourne le rotor.
Remarque : les sous-unités e, f, g, ATP8/A6L et j/6.8PL contribuent également au domaine membranaire de la tige périphérique et dans le complexe dimérique ( dimérisation de l'ATP synthase).
Structure et dimérisation de l'ATP synthase bovine
(Vidéo : Spikes et coll)
Dans la figure ci-dessus, chaque cercle représente l’état chimique du site catalytique dans chaque sous-unité β,
Les sous-unités α, β, γ, δ et ε du domaine catalytique F1 sont respectivement rouge, jaune, bleu, indigo et vert, avec la tige centrale, i.e. sous-unités γ, δ et ε attachées à l’anneau c8 (gris foncé) dans le domaine membranaire en contact avec la sous-unité a/ATP6 (bleu bleuet).
Les sous-unités de la tige périphérique (PS), OSCP, b, d et F6 sont respectivement bleu sarcelle, rose clair, orange et magenta, et la sous-unité ATP8/A6L est rouge brique.
Dans la région de l’interface monomère-monomère, les sous-unités e, f, g, j/6.8PL et k/DAPIT sont respectivement kaki, jaune paille, vert forêt, vert écume de mer et rose foncé. Les groupes de tête phosphate de la cardiolipine et du phosphatidylglycérol (PG) sont écarlates et les chaînes acyle grises.
entraîne la rotation de FO, i.e. rotation de l'anneau c et de la tige centrale liée, i.e. sous-unités γ, δ et ε,
fournit l'énergie nécessaire par la rotation de la sous-unité γ à l’intérieur de F1 qui conduit chaque paire αβ à passer par des conformations ouvertes (open), serrées (tight) et lâches (loose) qui entraînent la formation d’ATP dans les trois sites catalytiques β de F1.
3. Comme les pseudo-symétries sont différentes, 8 et 3, il existe un décalage de symétrie entre F1 et le domaine de l’anneau c8 que doit résoudre (Structure of the human ATP synthase 2023) :
Le couplage parfait est prédit à partir du nombre de copies des sous-unités β catalytiques et de celui des sous-unités c, i.e. par exemple, 8:3 dans les mitochondries de mammifères ou 10:3 chez la levure que nous prendrons comme exemple.
b. Avec ce rapport H+/ATP, l’énergie libre de la translocation des protons ($\Delta\mu_{H^+}$), multiplié par 8, est plus négative que l’énergie libre de l’hydrolyse de l’ATP ($\Delta\G_{ATP}$), multiplié par 3, et par conséquent, le moteur FO domine le moteur F1, le forçant à synthétiser l’ATP.
2. Toutefois, lorsque ce rapport s'inverse, F1 domine FO, l’hydrolyse séquentielle de l’ATP au niveau de chacune des trois paires αβ de F1 fait tourner la sous-unité γ dans la direction opposée, faisant pivoter l’anneau c porteur de protons contre la sous-unité a/ATP6 dans FO et pompant les protons à travers la membrane.
a. Chez les bactéries, les F-ATPases sont réversibles, i.e. peuvent aussi bien hydrolyser que synthétiser l'ATP.
b. Chez les eucaryotes, dans les mitochondries et les chloroplastes, elle peut s'inverser, mais dans des conditions exceptionnelles.
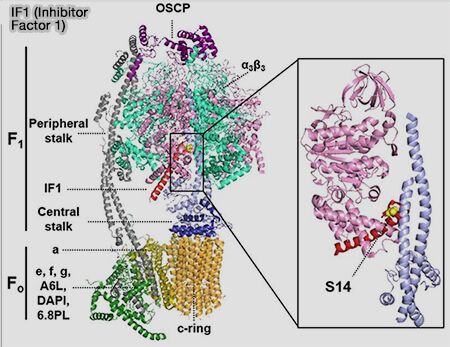
ATP synthase bovine et IF1
(Figure : vetopsy.fr d'après Domínguez-Zorita et coll)
Dans les mitochondries, l'activité normale implique un fort gradient d'H+ et un pH d'environ 7,8.
Lors d'ischémie, d'hypoxie ou d'apoptose, le gradient de protons (ΔpH) ou le potentiel de membrane (Δψ) s'effondrent, le pH diminue à environ 6,8 et l'ATP synthase peut fonctionner à l'envers pour pomper des protons ( chimiostose et phosphorylation oxydative).
La région N-terminale, i.e. 20-30 premiers acides aminés de I'F1 forme une hélice α qui s’insère dans une fente entre deux sous-unités β de F1, les deux domaines inhibiteurs faisant face à des sites opposés, interagissant ainsi simultanément avec deux domaines F1 adjacents.
Elle bloque mécaniquement le mouvement de rotation nécessaire à l’activité enzymatique.
Ce processus permet de protéger les cellules cardiaques, cérébrales et musculaires en cas d’hypoxie.
Remarque : dans les chloroplastes, la fonction de synthèse est aussi favorisée.





 dimérisation de l'ATP synthase).
dimérisation de l'ATP synthase).