Le mécanisme de la région F1 de l'ATP synthase, encore mal compris, est associé à des rotations de la tige centrale, conduisant à des changements conformationnels de l'enzyme liée à la production d'adénosine triphosphate (ATP).
Mécanisme de F1
Les mécanismes exacts ne sont toujours pas bien compris à l'heure actuelle.
Rotation des sous-unités (αβ)3
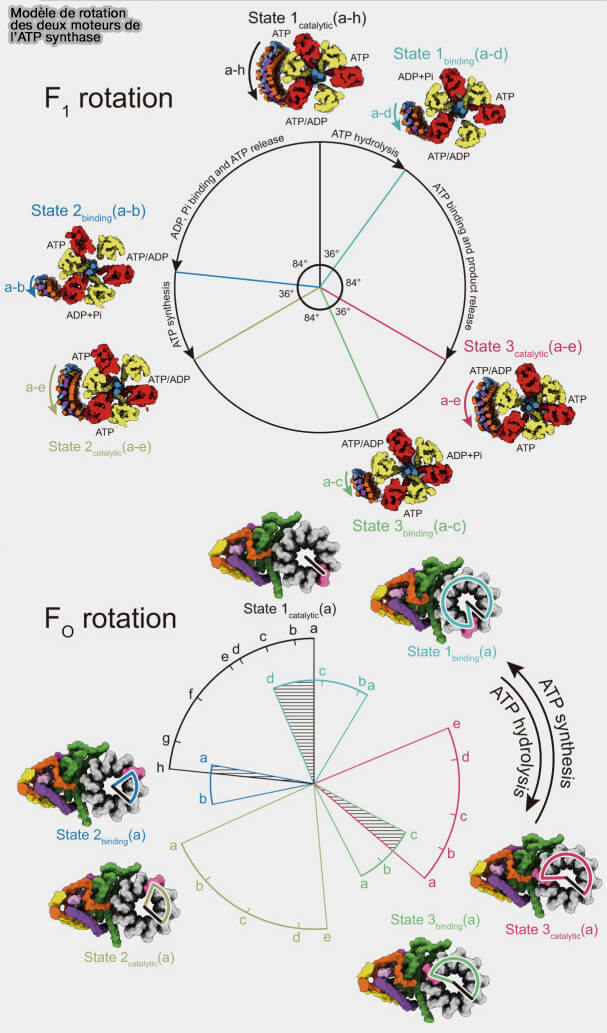
La synthèse, ou l'hydrolyse, de chaque molécule d'ATP est associée à une rotation de 120 ° de la sous-unité γ au sein de F1, conduisant à des conformations de l'enzyme connues sous le nom d'état rotationnel 1, 2 et 3, maintenant divisé en 6 stades.
Les sous-unités α et β sont alternées et forment un anneau globulaire d'environ 100 Å de diamètre, i.e. (αβ)3, avec 6 sites de liaison aux nucléotides.
1. Les sites de liaison aux nucléotides, sous-unités (αβ)3, sont contenus dans :
dans les trois sous-unités α catalytiquement inactives, i.e. αE, αDP et αTP,
dans les trois unités β qui catalysent la synthèse de l'ATP, i.e . βE, βDP et βTP.
Remarque : les articles diffèrent selon les nucléotides liés aux sous-unités α.
2. Les trois unités β subissent des changements conformationnels qui entraînent la rotation de la sous-unité γ pendant l'hydrolyse de l'ATP.
a. Les trois unités β comprennent :
βE, à conformation ouverte, qui est vide, i.e. empty en anglais, i.e. 0° sur la figure ci-dessous,
βTP, à conformation serrée (tight), qui contient un MgATP, i.e. rotation à 120°,
βDP, à conformation lâche (loose), qui contient un MgADP et un Pi, i.e. rotation à 240°.
Les trois sites catalytiques présentent des différences marquées dans leur affinité pour le Mg-nucléotide, i.e. élevée pour βTP, moyenne pour βDP et faible pour βE, qui sont déterminées par la position de la sous-unité γ centrale, i.e. après rotation de y de 120 °, les sites ont échangé leurs affinités.
La position 80 °, appelée “ binding dwell ”, i.e. dwell, demeurer en anglais, correspond à l'étape d'attente de la liaison de l'ATP ou à la libération du phosphate (Pi), i.e. attente de l'hydrolyse.
La position 40°, appelée “ catalytic dwell ” semblait être une étape d'attente de l'hydrolyse, qui correspondrait, selon les auteurs, à l'hydrolyse effective de l'ATP ou à la libération de l'ADP.
4. Dans la synthèse de l'ATP, la position 80 ° correspond à l'étape d'attente de la liaison de l'ADP et du Pi, la position 40 ° à celle de la synthèse effective de l'ATP.
Interaction des sous-unités (αβ)3 avec la sous-unité γ
À l'heure actuelle, la connaissance des interactions entre les sous-unités γ et β (et/ou α) qui sont responsables des différentes affinités des sites de liaison catalytique est fragmentaire.
Les différences et les changements d'affinité semblent être d'importance capitale pour le mécanisme enzymatique en ce qui concerne le couplage entre la rotation et la catalyse ainsi que la catalyse elle-même.
1. La synthèse et l'hydrolyse de l'ATP ne se produisent que sur le site de haute affinité.
Dans l'hydrolyse de l'ATP, la fermeture du site de faible affinité autour de l'ATP nouvellement lié, accompagnée d'une conversion du site à forte affinité, est largement considérée comme entraînée par la rotation à 80 °.
b. L'hélice C-terminale de γ, i.e. résidus 256-265, approche de la boucle de capture, i.e. catch loop, de la βE, i.e. résidus 309-316.
L'absence de ce segment inhibe cette interaction.
Sous-unité γ et différents sites catalytiques
(Figure : vetopsy.fr d'après Mnatsakanyan et coll)
Rôle de la sous-unité δ/ε
La sous-unité δ au niveau mitochondrial, qui correspond à la sous-unité ε au niveau des bactéries et des chloroplastes, inhiberait la rotation de la sous-unité γ.
1. Il existe une inadéquation entre les rotations des domaines catalytiques αβ de pseudo-symétrie 3, et l'anneau c, de pseudo-symétrie 8 ou plus, que doivent résoudre (Structure of the human ATP synthase 2023) :
Les mécanismes exacts ne sont toujours pas bien compris à l'heure actuelle.
2. La tige périphérique (PS) est un composant essentiel du stator de l'enzyme reliant (αβ)3 du domaine catalytique F1 à la région en forme de coin associée à la membrane et à la sous-unité a/ATP6 adjacente dans le sous-complexe du stator contre lequel le rotor tourne.
elle empêche la dissociation du domaine (αβ)3 de la tige centrale en la plaquant à l'aide d'OSCP,
elle résiste au couple de rotation de la tige centrale, empêchant le domaine (αβ)3 et le reste du domaine du stator de suivre la direction de la rotation.
Cette vidéo montre les mouvements latéraux étendus de la tige périphérique et le mouvement de bascule du domaine catalytique alors que la tige centrale asymétrique tourne dans le domaine F1.
Les résidus 1-25 des sous-unités α sont rouge pâle.
Les sous-unités ATP8/A6l, a, b, d, F6 et l'OSCP sont respectivement rouge brique, bleu bleuet), rose clair, orange, magenta et bleu sarcelle.
La coiffe, i.e. cap en anglais, est composée par le domaine N-terminal de l'OSCP.
La charnière, reliant la coiffe au domaine C-terminal de l'OSCP, qui permet à la tige périphérique de s'adapter aux mouvements verticaux et transversaux du domaine (αβ)3 pendant la catalyse causée par le rotor asymétrique.
La tige rigide de 150 Å de longueur est formée par des portions parallèles de régions α-hélicoïdale des sous-unités b, d, F6 et la région C-terminale de ATP8/A6L.
La charnière de la sous-unité b s'attache à la sous-unité a/ATP6 de FO.
3. Ainsi, les déplacements latéraux de la tige périphérique provoqués par la rotation de la tige centrale asymétrique sont transmis au domaine membranaire, avec une réduction du déplacement du domaine catalytique, i.e. processus qui pourrait augmenter l'efficacité de l'enzyme.
Un certain degré de mouvement du domaine catalytique doit être autorisé pour permettre une conversion efficace du couple de rotation de la tige centrale en changements conformationnels dans les sous-unités catalytiques, mais sans gaspiller de l'énergie par déplacement inutile de la partie extrinsèque de l'enzyme.
La flexibilité de la tige périphérique mène efficacement cet équilibre, empêchant la rotation libre du domaine catalytique, pour s'adapter aux mouvements de balancement induits par la tige centrale asymétrique et le déplacement d'amortissement du domaine membranaire aidant à maintenir la sous-unité a en place.
Chez E. Coli ou chez la levure
La tige périphérique (PS) de l’ATP synthase d’Escherichia coli ou de S. cerevisiae est plus simple et consiste en un domaine coiled-coil (superhélice) α-hélicoïdal formé par les deux sous-unités b.








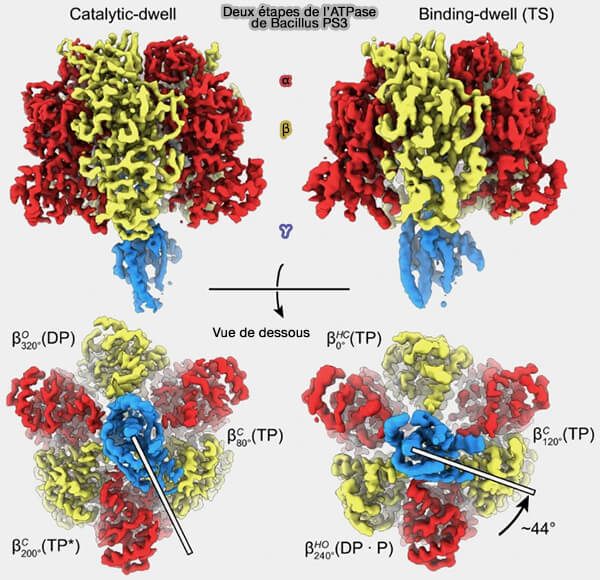
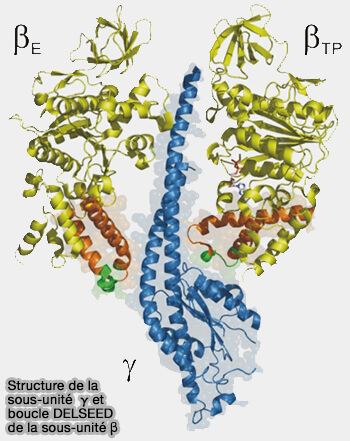
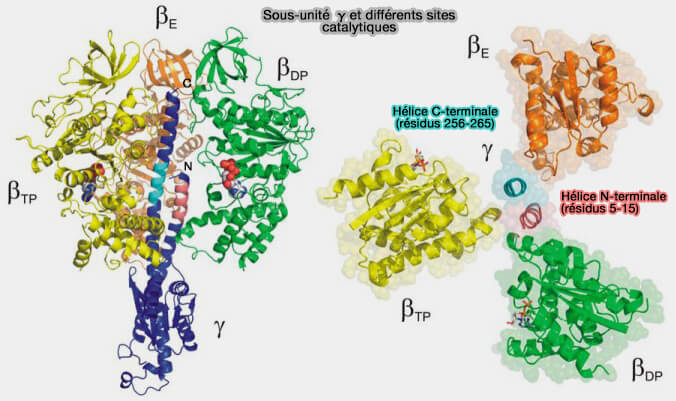
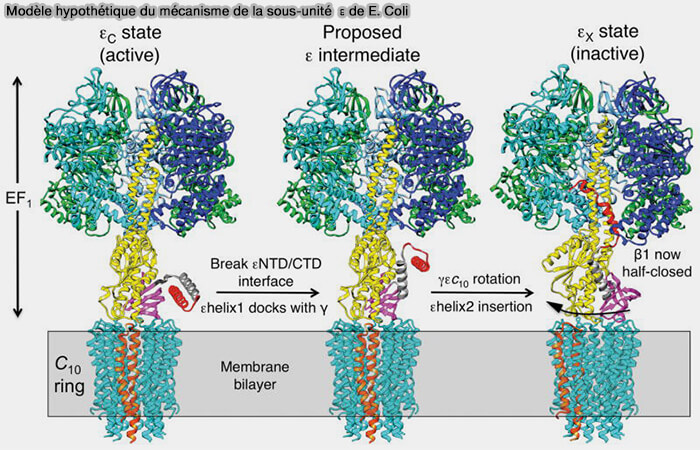
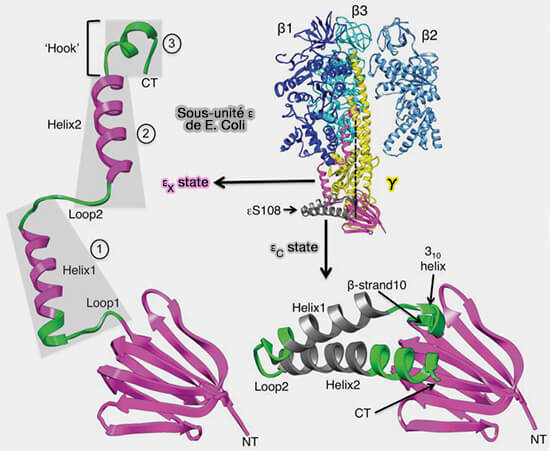
 figure S23) :
figure S23) :