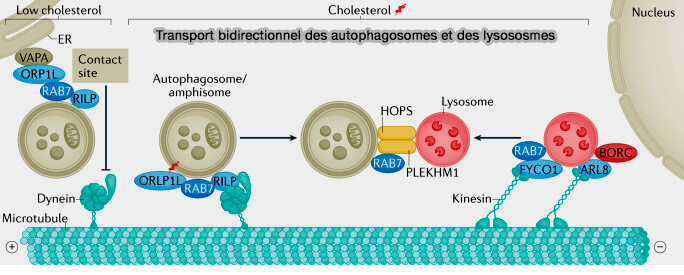
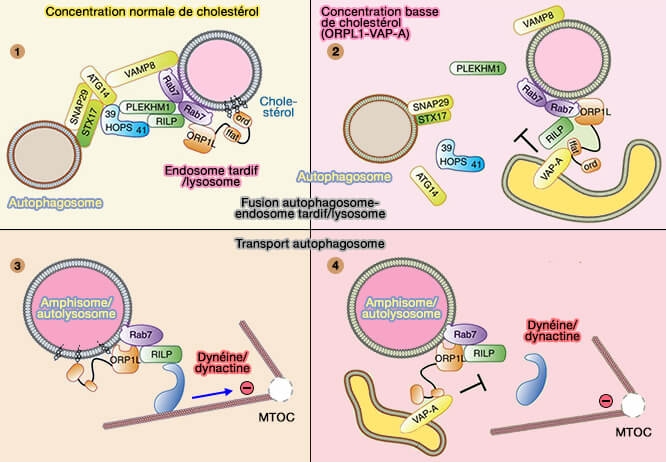
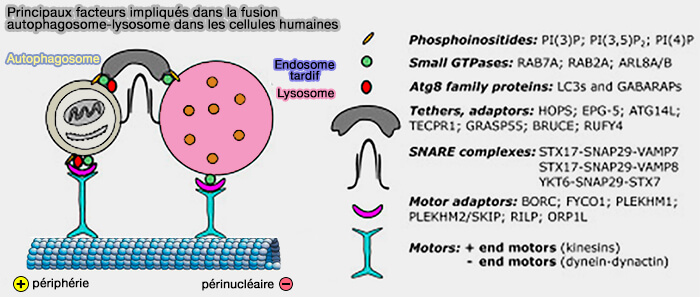
Les autophagosomes et les endosomes tardifs (LE)/lysosomes subissent un mouvement bidirectionnel sur les microtubules pour pouvoir fusionner.
Principaux facteurs impliqués dans la fusion des autophagosomes
(Figure : vetopsy.fr d'après Lőrincz et Guhász)
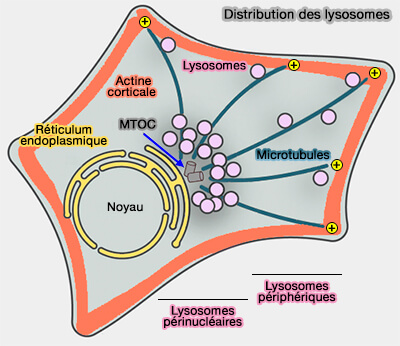
Vue d'ensemble des déplacements des autophagosomes
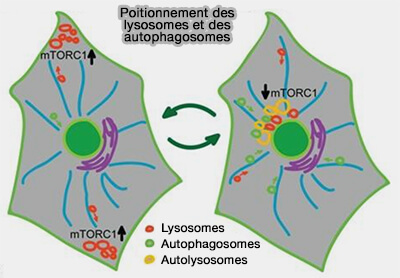
Les autophagosomes sont générés à des positions aléatoires dans le cytoplasme et les lysosomes sont localisés principalement dans la région périnucléaire.
Positionnement des lysosomes et des autophagosomes
(Figure : vetopsy.fr d'après Korolchuk et coll)
1. Les autophagosomes et les endosomes tardifs (LE)/lysosomes subissent un mouvement bidirectionnel sur les microtubules.
Les LE/lysosomes recrutent des kinésines et des adaptateurs pour leur transport antérograde ( trafic antérograde).
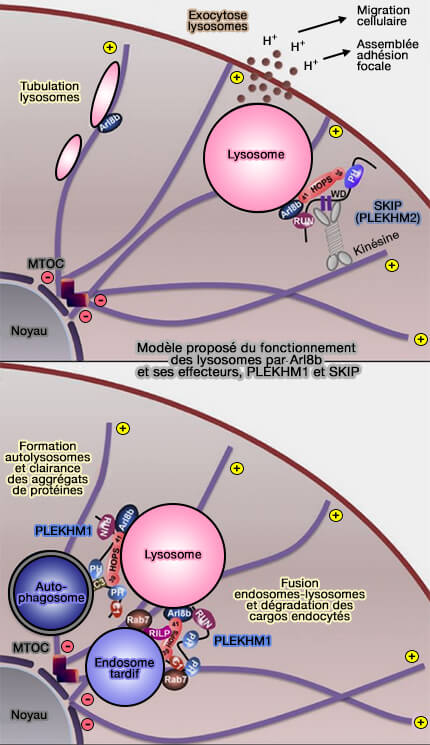
Les protéines effectrices des petites GTPases Rab et Arf-like, i.e. Arl8, comprennent des adaptateurs de liaison aux protéines motrices, qui sont capables de conduire le transport des autophagosomes et des endosomes/lysosomes tardifs les uns vers les autres le long du cytosquelette.
l'augmentation des taux de fusion, tandis que la surexpression de la kinésine-3 redistribuait les autophagosomes vers la périphérie et que le taux de leur fusion avec les lysosomes diminuait considérablement.
Ces résultats suggèrent que le positionnement des lysosomes est capable de déterminer le taux de fusion autophagosome/lysosome.
2. Le transport centripète dépend de RILP, effecteur de Rab7, qui interagit avec le complexe de dynéine-dynactine.
2. ARL8 relie également les lysosomes à la dynéine motrice par RUFY3 et RUFY4, fournissant des informations clés sur le mouvement bidirectionnel des lysosomes.
la sortie initiale des autophagosomes de l’axone distal,
le transport rétrograde soutenu le long de l’axone moyen,
la maturation autophagosomale dans l’axone proximal.
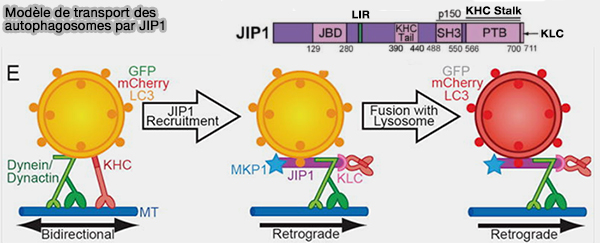
2. JIP1 se lie directement à la dynactine, l’activateur de la dynéine, mais se lie également à la chaîne lourde (KHC) kinésine-1 et l’active de manière dépendante de la phosphorylation sur S421.
L'inhibition de cette phosphorylation pour conserver un transport rétrograde s'effectue par :
les phosphatases MKP1 autophagosomales,
la liaison de LC3.
Modèle de transport des autophagosomes par JIP1
(Figure : vetopsy.fr d'après Fu et coll)
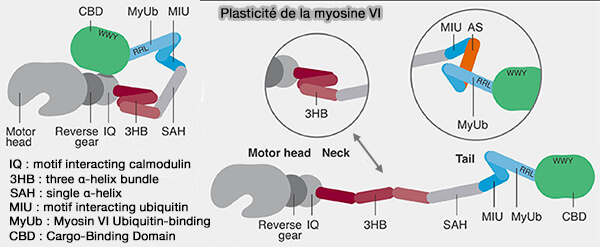
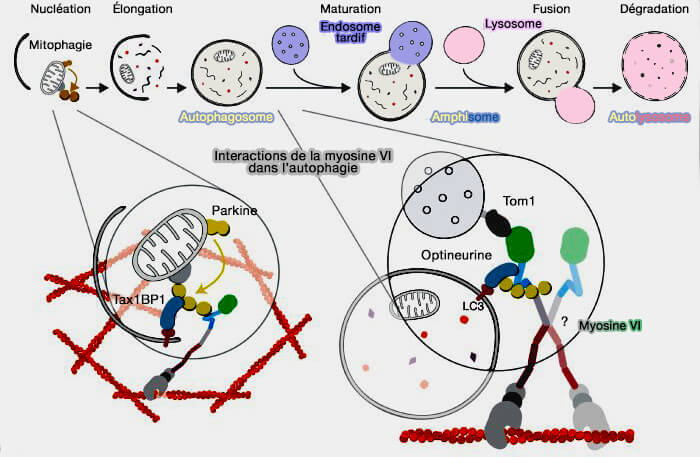
a. La myosine VI est le seul moteur à fonctionner à contre-sens, i.e. vers l'extrémité (-)
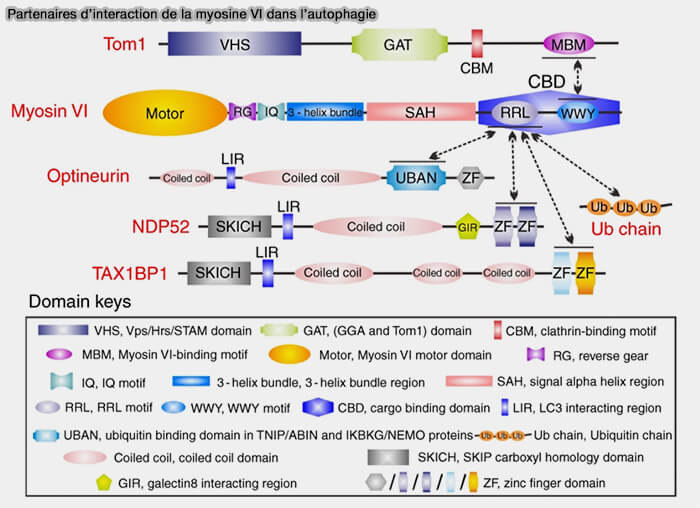
b. Les adaptateurs MYO6 et les récepteurs de l’autophagie peuvent former le pont entre les bactéries ou les mitochondries ubiquitinées et LC3 sur la membrane du phagophore comme :
Partenaires d’interaction de la myosine VI dans l’autophagie
(Figure : vetopsy.fr d'après Hu et coll)
4. À des stades ultérieurs, l’ubiquitination des récepteurs de l’autophagie favorise la fusion des autophagosomes avec les endosomes par la myosine VI qui se lie simultanément (Myomics: myosin VI structural and functional plasticity 2021) :




 trafic antérograde).
trafic antérograde).