


Autophagie
Protéines ATG (AuTophaGy-related)
Atg9
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

ATG9/Atg9 est la seule protéine transmembranaire dans la machinerie autophagique et est essentielle à l’initiation de la formation des autophagosomes.
Structure d'ATG9
Atg9 est une scramblase transmembranaire à passage multiple dont la structure n'a pas encore été complètement mise à jour.

Vous pouvez lire : Structure of Human ATG9A, the Only Transmembrane Protein of the Core Autophagy Machinery (2020) et Subnanometer resolution cryo-EM structure of Arabidopsis thaliana ATG9 (2020).
Les deux structures, humaine et végétale, diffèrent par quelques points.
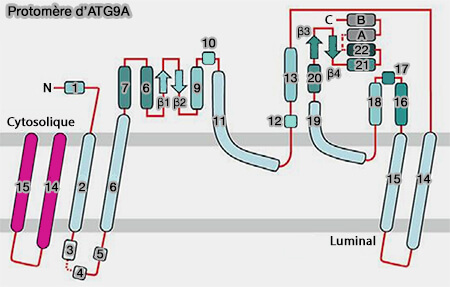
- Protomère d'ATG9A
(Figure : vetopsy.fr d'après Guardia et coll)
1. Atg9 comprend :
a. des domaines transmembranaires :
- 4 domaines transmembranaires pour ATG9A, i.e. α2, α6, α14 et α15 avec deux hélices qui sont partiellement encastrées dans la membrane, i.e. α11 et α19, à
- 6 domaines transmembranaires pour Atg9,
b. les extémités N- et C-terminales cytosoliques.
Remarque : α6, inhabituellement longue, s'étend dans le cytosol, formant ainsi une ancre structurelle qui entre en contact avec différentes parties au sein du protomère.
2. ATG9A se présente sous forme de trimère au niveau membranaire, avec deux conformations, A et B, dans lesquelles les protomères ont été tournés de 8 degrés ( vidéo).
vidéo).
Les interactions entre les domaines des interfaces sont complexes (revue dans les articles précédents).
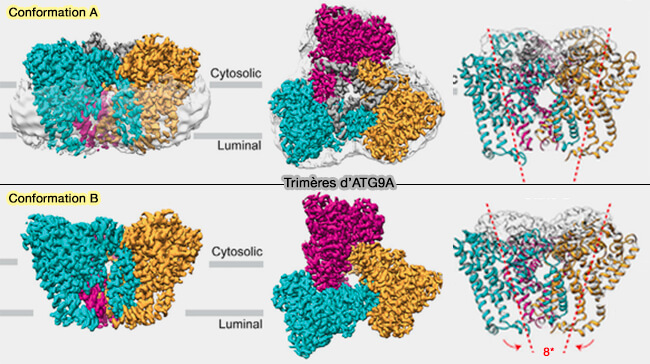
(Figure : vetopsy.fr d'après Guardia et coll)
a. Ce trimère présente un pore central qui se ramifie en :
- un pore perpendiculaire,
- un pore horizontal.
b. ATG9A comprend ainsi un réseau arborescent de pores avec une seule ouverture du côté luminal et plusieurs ouvertures du côté cytoplasmique.
c. Les deux conformations diffèrent par l'ouverture du pore central, expliquant le passage des phospholipides lors de la formation de l'autophagosome.
Les deux conformations ont une affinité pour les membranes incurvées, i.e. les vésicules d'ATG9.
Rôle d'ATG9
1. Dans des conditions riches en nutriments, l'ATG9A est principalement située dans la région périnucléaire :
- au niveau l'appareil de Golgi,
- mais aussi, moins fortement, au niveau des endosomes précoces et de recyclage (Atg9A trafficking through the recycling endosomes 2016).
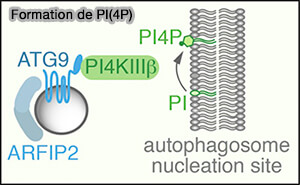
(Figure : vetopsy.fr d'après Judith et coll)
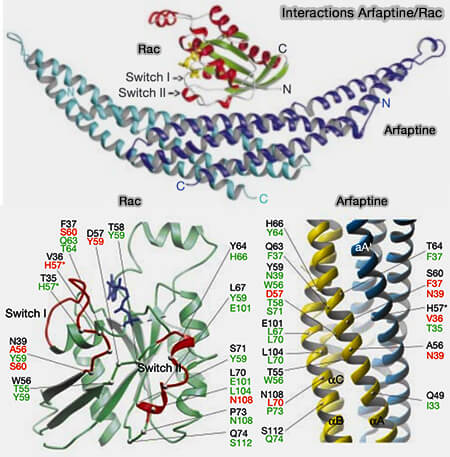
2. Pendant la dénutrition, l'ATG9A périnucléaire diminue concomitamment avec une augmentation d'une population vésiculaire de 30–60 nm riche en arfaptines et en PI4-kinases :
- PI4KIIIβ, kinase qui produit du PI(4)P, au niveau des omégasomes,
- PI4KIIα, qui intervient dans la fusion autophagosome/lysosome (GABARAPs regulate PI4P-dependent autophagosome:lysosome fusion 2015).
a. Les vésicules ATG9A transportent PI4KIIIβ vers la membrane du réticulum endoplasmique (RE), favorisant la production de PI(4)P, sur le site d'initiation, ce qui facilite le recrutement du complexe ULK et l'initiation de l'autophagie (ATG9A shapes the forming autophagosome through Arfaptin 2 and phosphatidylinositol 4-kinase IIIβ 2019),
b. PI4KIIα intervient plus tardivement dans la fusion autophagosome/lysosome (GABARAPs regulate PI4P-dependent autophagosome:lysosome fusion 2015).

Le rôle clé de l'ATG9A dans l'autophagie peut être de fournir PI(4)P aux membranes autophagosomales.
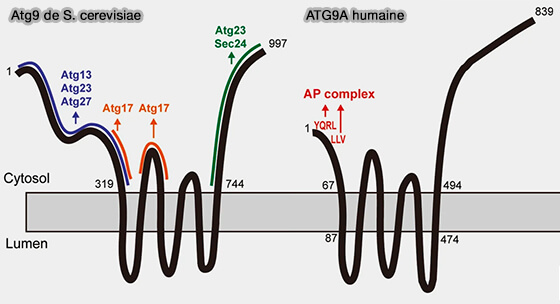
3. Toutefois, il existe une différence majeure entre l'Atg9 de la levure et l'ATG9 des mammifères.
a. Chez la levure, un modèle de la formation du phagophore a été proposé en considérant que les vésicules d'ATG servent de plateforme à la machinerie autophagique (The Role of ATG9 Vesicles in Autophagosome Biogenesis 2024).
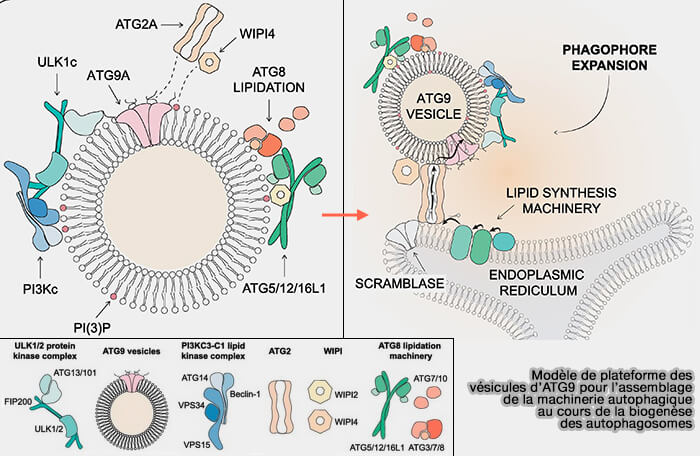
(Figure : vetopsy.fr d'après Holzer et coll)
b. Chez les mammifères, les vésicules interagiraient de manière transitoire et ne seraient pas incorporées dans un autophagosome complet (Autophagy initiation by ULK complex assembly on ER tubulovesicular regions marked by ATG9 vesicles 2016).
- L'ATG9A fournit des composants clés, tels que des protéines et des lipides, aux membranes autophagosomales.
- Malgré les différences apparentes dans la localisation de l'ATG9A lors de l'initiation de l'autophagie, les deux modèles proposés dans les cellules de levure et de mammifère ne s'excluent pas mutuellement.
Remarque : une étude récente suggère qu'ATG9 se lierait aussi directement à LC3 dans les phagophores et aurait la capacité de promouvoir la formation des autophagosomes (Mammalian ATG9s drive the autophagosome formation by binding to LC3 2020).
Distribution d'ATG9
Vue d'ensemble
Chez la levure
Chez la levure, la distribution d'Atg9 est en partie régulée par le complexe Atg1 via ses interactions physiques avec d'autres protéines Atg du complexe.
1. Les régions cytoplasmiques d'Atg9 interagissent avec l'Atg17 (The Atg1–kinase complex tethers Atg9-vesicles to initiate autophagy 2016).
a. Atg17 forme des dimères anti-parallèles qui attachent les vésicules Atg9 indépendamment des autres cofacteurs, mais cette activité est inhibée par les sous-unités Atg31-Atg29 (Structural characterization of the Saccharomyces cerevisiae autophagy regulatory complex Atg17-Atg31-Atg29 2018).
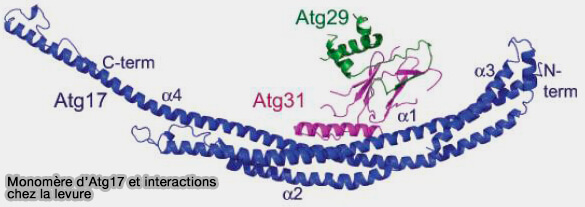
(Figure : vetopsy.fr d'après Ragusa et coll)
- L'extrémité N-terminale d'Atg9 se lie également au domaine HORMA d'Atg13, une autre sous-unité du complexe Atg1, ce qui facilite le recrutement des vésicules Atg9 au PAS (Atg13 HORMA domain recruits Atg9 vesicles during autophagosome formation 2015).
(Figure : vetopsy.fr d'après Rao et coll)
Remarque : Atg17 intervient dans l'autophagie non sélective tandis qu'Atg11 est impliquée dans l'autophagie sélective ( autophagie sélective chez la levure).
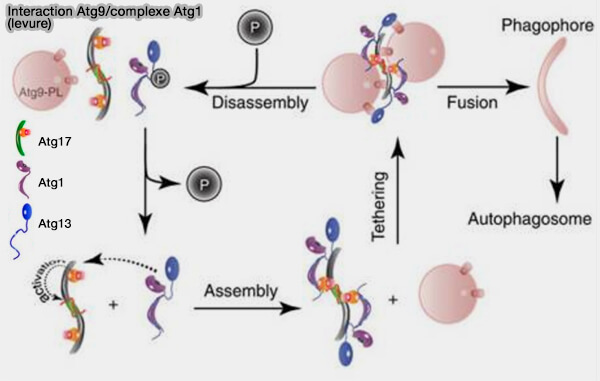
b. Atg9 est phosphorylé par la kinase Atg1, et cette phosphorylation est nécessaire pour le recrutement de facteurs en aval tels qu'Atg18/WIPI et Atg2, alors qu'elle n'affecte pas le recrutement PAS d'Atg9 (Early Steps in Autophagy Depend on Direct Phosphorylation of Atg9 by the Atg1 Kinase 2014).
(Figure : vetopsy.fr d'après Nishimura et Tooze)
Remarque : si Atg17 intervient dans la macroautophagie non sélective, Atg17 est impliquée dans l'autophagie sélective.
2. Chez les mammifères, comme chez la levure, le trafic d'ATG9A dépend du complexe ULK, bien qu'une analyse plus approfondie soit nécessaire pour confirmer si ATG9A interagit directement avec lui (Regulation of mATG9 trafficking by Src- and ULK1-mediated phosphorylation in basal and starvation-induced autophagy 2016).
Trafic d'ATG9
1. PI4KIIIβ interagit avec ATG9A pour contrôler la production de PI(4)P au site d’initiation de la membrane et la réponse autophagique (remplacement de PI(3)P par PI(4)P).
- PI4KIIIβ et PI(4)P fonctionnent probablement en recrutant la sous-unité ATG13 du complexe ULK dans les autophagosomes naissants.
- Le trafic d'ATG9A à partir du complexe de l'appareil de Golgi vers les omégasomes est contrôlé positivement par le complexe ULK1/2 et négativement par le complexe mTORC1.
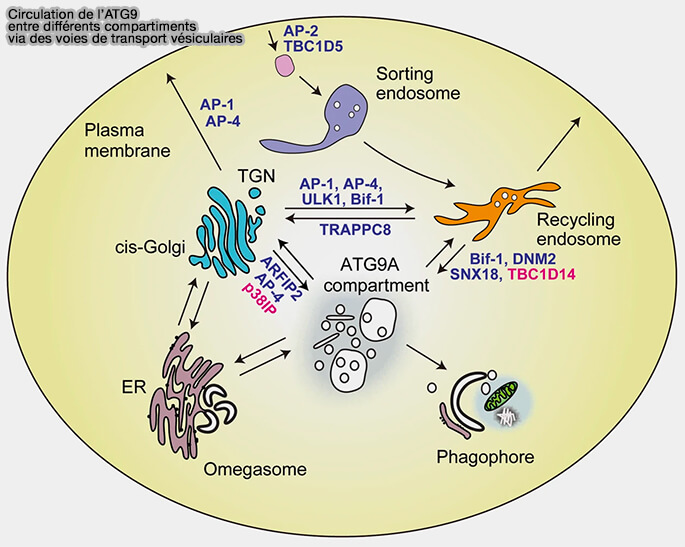
(Figure : vetopsy.fr d'après Nishimura et Tooze)
1. Le trafic d'ATG9A à partir du Golgi nécessite également l'endophiline B1 (BIF-1) en coopération avec la dynamine pour la formation des vésicules ATG9A à partir des endosomes de recyclage (The Bif-1-Dynamin 2 membrane fission machinery regulates Atg9-containing vesicle generation at the Rab11-positive reservoirs, 2016).
- Il semblerait que la sphingomyéline phosphodiestérase 1 (SMPD1) contrôle ce trafic (Excess sphingomyelin disturbs ATG9A trafficking and autophagosome closure 2016).
- D'autres protéines contribuent à la sortie ATG9A des endosomes de recyclage, notamment P38IP, (p38 MAPK-interacting protein) et la nexine 18 (SNX18 regulates ATG9A trafficking from recycling endosomes by recruiting Dynamin‐2 2018).
- PI(4)P) peut également réguler le trafic d'Atg9, car PI4KIIIβ est essentiel pour la sortie d'Atg9 du Golgi (Phosphatidylinositol 4-Kinases Are Required for Autophagic Membrane Trafficking 2012).
2. Les adaptateurs AP-1, AP-2 et AP-4 régulent également le trafic d'ATG9A par leur interaction avec son extémité N-terminale.
- AP-2 affecte probablement le transport de l'ATG9A à partir de la membrane plasmique et des endosomes de recyclage (Regulation of mATG9 trafficking by Src- and ULK1-mediated phosphorylation in basal and starvation-induced autophagy 2017
- AP-4 régule la sortie du réseau trans-Golgi ou TGN (AP-4 mediates export of ATG9A from the trans-Golgi network to promote autophagosome formation 2017).
Remarque : Atg9 de la levure n'a pas de signaux de reconnaissance complexes d'adaptateurs, mais Atg23 et Atg27 exportent Atg9 dans les vésicules dérivées du Golgi.
3. Rab1B/Rab11 contrôle le trafic entre :
- le réticulum endoplasmique (RE) et le Golgi (WYpt1/Rab1 regulates Hrr25/CK1δ kinase activity in ER–Golgi traffic and macroautophagy 2015) l
- le Golgi et les endosomes de recyclage (The Function of the Intermediate Compartment in Pre-Golgi Trafficking Involves its Stable Connection with the Centrosome 2009).
(Figure : vetopsy.fr d'après Tarricone et coll)
Rab1, i.e. Ypt1 de la levure, contrôle le complexe TRAPP-III qui régule le trafic ATG9A/Atg9.
4. L'arfaptine-2 (ARFIP2), protéine à domaine BAR, servirait de régulateur pour la détection et la génération de courbure membranaire et pour sa liaison avec Rab1/11.
Les ARFIP contiennent une hélice amphipathique (AH), qui confère une spécificité pour se lier aux liposomes contenant du PI(4)P) et au TGN (Recruitment of arfaptins to the trans-Golgi network by PI(4)P and their involvement in cargo export 2013).
Interactions avec Atg2 dans la formation des autophagosomes
Vous pouvez lire, pour plus de détails : ATG9A and ATG2A form a heteromeric complex essential for autophagosome formation (2022).
1. Des protéines à domaine choréine telles que ATG2 ou VPS13, protéines à domaine choréine, peuvent relier deux membranes par un long pont protéique de 16 nm en forme de tunnel avec une rainure hydrophobe à travers laquelle les lipides peuvent se déplacer, alors que la protéine reste stationnaire (ATG2 and VPS13 proteins: molecular highways transporting lipids to drive membrane expansion and organelle communication 2021).
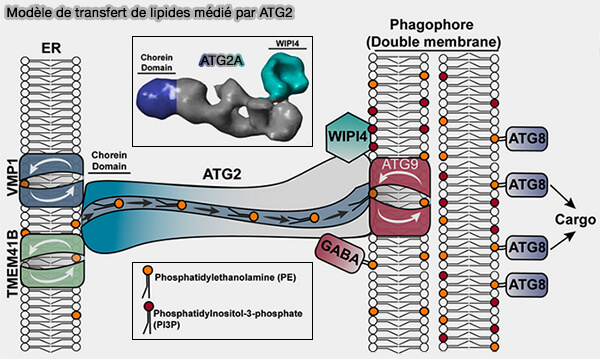
(Figure : vetopsy.fr d'après MacEwan et coll)
2. Atg9, forme formant un complexe hétérotétramérique avec Atg2, i.e. domaines C-terminal du trimère d'Atg9 et domaine N-terminal d'Atg2.
Le modèle suppose qu'une fois que les lipides sont liés à la branche perpendiculaire de l'ATG9A, ils peuvent accéder à la membrane par le pore central ou la branche latérale dans chaque protomère.
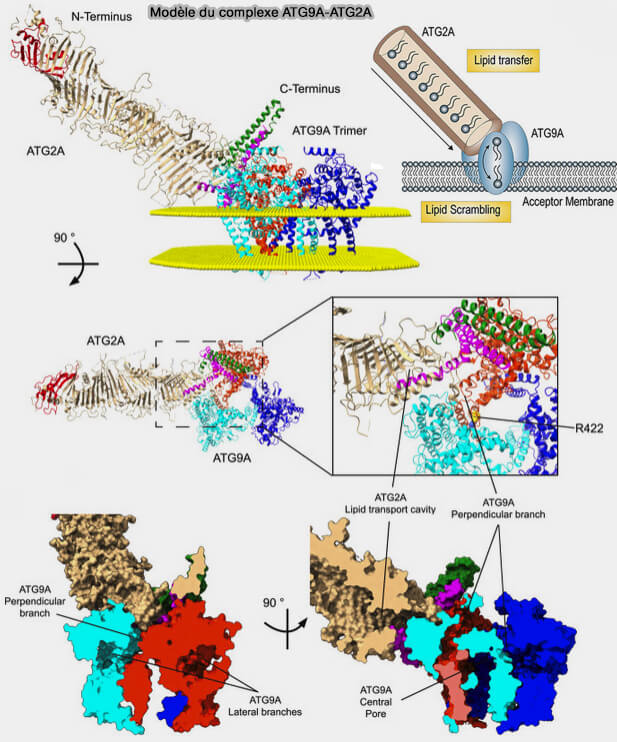
(Figure : vetopsy.fr d'après Van Vliet et coll)
Retour à l'initiation du phagophore
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation