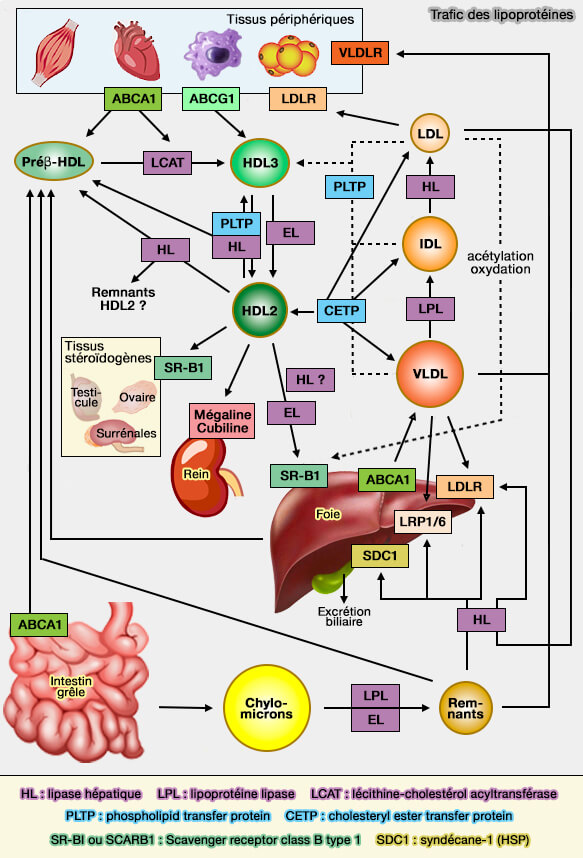
Modèle de transfert " navette " de CETP
(Figure : vetopsy.fr d'après Shrestra et coll)
1. Ce processus conduit à la formation de complexes binaires qui facilitent les échanges bidirectionnels d'esters de cholestérol (CE) et de triglycérides (TG) entre chacune des lipoprotéines et CETP.
Les complexes se dissocient par la suite des lipoprotéines où ils ont été générés, et restent dans la circulation jusqu'à ce qu'ils entrent au hasard avec une autre lipoprotéine et participent à un autre cycle d'échanges CE et TG.
De plus, la CETP transfère les lipides via les propriétés de son domaine C-terminal, qui ordonne les lipides via des transitions désordonnés à ordonnés dans sa structure secondaire.
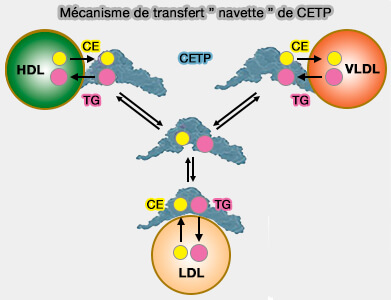
a. Les groupes phosphate des phospholipides sont les principaux sites d'interaction des lipoprotéines avec la CETP, en particulier avec les lysines de la surface concave.
b. Des résidus glutamate et aspartate forment des ponts salins avec les groupes choline chargés positivement des POPC, bien que le rapport des ponts salins formés par les acides aminés chargés positivement et négativement soit d'environ 1,8, ce qui indique que la plupart des acides aminés chargés positivement contribuent à la formation de complexes CETP-lipoprotéine.
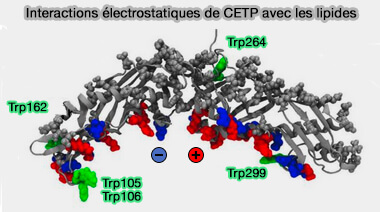
La région Ω5 du volet flexible de CETP et les résidus Trp qui s'y trouvent est cruciale dans la liaison de la CETP aux particules HDL.
4. La profondeur de pénétration de la CETP est un facteur important dans l'échange de lipides médié par la CETP, car elle détermine l'efficacité avec laquelle les lipides du noyau neutre peuvent interagir avec la CETP.
La CETP se fixe à la surface à l'aide d'interactions électrostatiques, les groupes de tête des POPC s'écartent, donnant accès à la région lipidique hydrophobe.
Par ce phénomène, les deux ouvertures du tunnel de la surface concave sont exposées à la matrice lipidique hydrophobe de la lipoprotéine.
La formation de patch hydrophobe facilite, en générant un défaut à la monocouche de surface, la diffusion des lipides centraux vers la région de la monocouche de surface située sous CETP.
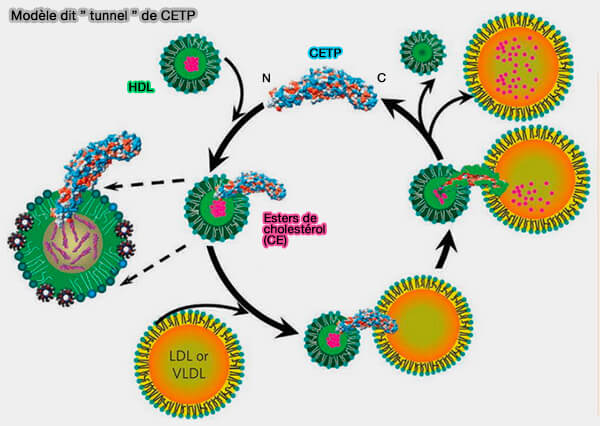
Modèle dit " tunnel "
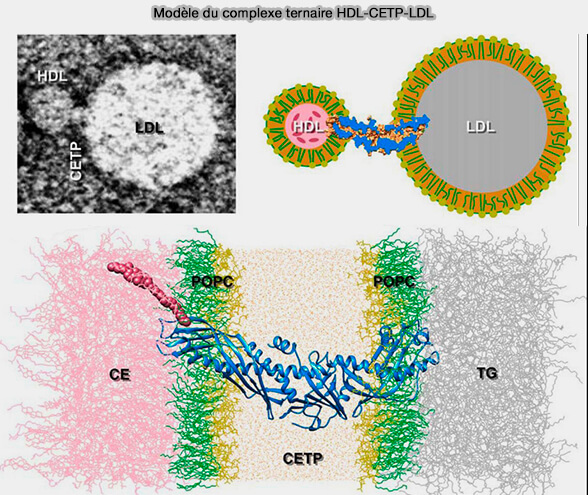
Le deuxième modèle, dit " tunnel " implique la formation d'un pont entre la CETP et deux particules de lipoprotéines.
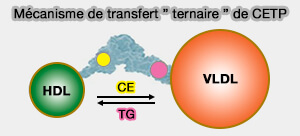
Modèle de transfert " ternaire " de CETP
(Figure : vetopsy.fr d'après Shrestra et coll)
Le domaine N-terminal de CETP pénétrant dans la monocouche de surface HDL-phospholipide à une profondeur d'environ 17 à 28 Å et atteignant le noyau HDL-CE.
Le domaine C-terminal uniquement pénètre la surface des LDL ou des VLDL à une profondeur de 20 à 25 Å.
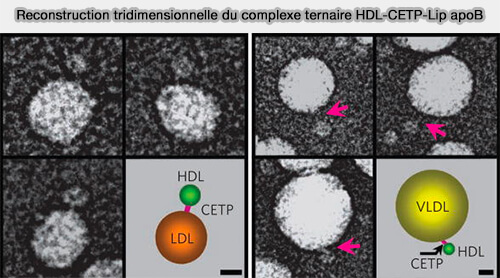
Reconstruction tridimensionnelle du complexe HDL-CETP-Lip apoB
(Figure : vetopsy.fr d'après Zhang et coll)
a. L'apoA-I et l'apoA-II peuvent ne pas être impliquées dans la liaison de la CETP.
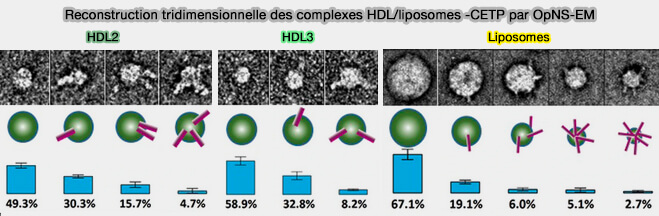
Reconstruction tridimensionnelle du complexe HDL-liposomes-CETP
(Figure : vetopsy.fr d'après Zhang et coll)
b. Les phospholipides (PL) de surface et la courbure de surface dominent probablement l'interaction de CETP avec la surface de HDL. Une plus grande surface lipidique offre une plus grande possibilité de liaison pour CETP.
HDL2, dont le diamètre est supérieur à celui de HDL3, a également un pourcentage plus élevé de particules CETP liées (50% contre 42%). Les lipides de surface de HDL2 occupent une plus grande surface que les lipides de surface de HDL3 (31,3% contre 22,7%).
Les liposomes plus gros que HDL2 ou HDL3 possèdent environ 6 molécules de CETP liées, par rapport à 1 ou 2 molécules de CETP pour HDL2 et HDL3.
Ce pore pourrait être important pour le transfert de phospholipides (PL) de surface lorsque la taille des HDL diminue, par exemple lors de l'efflux de cholestérol, stabilisant ainsi potentiellement la courbure croissante ou décroissante de la surface des HDL.
Modèle dit " tunnel " de CETP
(Figure : vetopsy.fr d'après Zhang et coll)
Dans le modèle clef-verrou, l'hélice X est une porte pour le transfert de lipides avec des états ouverts et fermés, i.e. les CE diffuseraient dans le tunnel hydrophobe du CETP lorsque la conformation de l'hélice X serait à l'état ouvert ou lorsqu'elle se déplace à l'intérieur du tunnel hydrophobe.
Dans l'autre modèle, le domaine du tonneau β N-terminal de la CETP agirait comme une porte pour le transfert des lipides et l'hélice X est présente et principalement dans son état fermé, ce qui est très probablement une condition nécessaire au bon fonctionnement du transfert des lipides.
Il semblerait que ces résultats contardictoires soient dus aux anticorps utilisés pour cibler les domaines N- et C-terminaux de la molécule CETP.
Remarque : le modèle tunnel dimère, un modèle tunnel modifié, implique la formation d'un tunnel hydrophobe par un dimère CETP au lieu d'un monomère.
Bien que tous ces modèles rendent compte du processus de transport lipidique de base, le mécanisme détaillé est inconnu en raison des propriétés dynamiques et des flexibilités structurelles des lipoprotéines et des complexes CETP-lipoprotéine.