SR-B1 (Scavenger Receptor Class B type 1 ou SCARB1) est une protéine réceptrice membranaire qui fonctionne comme un récepteur des HDL (lipoprotéines de haute densité) pour la livraison du cholestérol aux cellules.
mais aussi, dans une moindre mesure, les adipocytes, les macrophages, les cellules endothéliales, les cellules musculaires lisses, les cellules intestinales, les cellules rétiniennes et les kératinocytes cutanés.
2. SR-B1, par son domaine extracellulaire, se lie à de nombreux ligands :
a. Comme la structure à haute résolution n'a pas encore été révélée, les chercheurs se sont appuyés sur des études de mutagenèse et sur la résolution des structures de CD36 ou LIMP-2 :
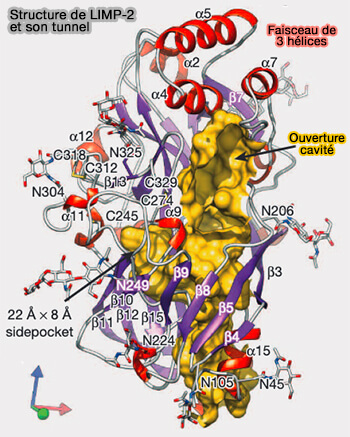
Structure de LIMP-2 et son tunnel
(Figure : vetopsy.fr d'après Neculai et coll)
b. Un modèle d'homologie de SR-B1 humain pleine longueur a été créé à l'aide de trRosetta, i.e. un algorythme pour la prédiction des structures protéiques
Le domaine extracellulaire (ECD) serait étendu, i.e. résidus 36-439.
1. Le domaine extracellulaire est composé d'un tonneau β antiparallèle central et des hélices α courtes périphériques, i.e. structure entrelacées reliées par des boucles qui confèrent probablement une flexibilité conformationnelle.
a. Un faisceau tri-hélicoïdal à l'apex est composé des résidus A144-N150, M153-L166 et P186-Y194.
Les mutations M159E et T165E au sein de ce faisceau hélicoïdal de SR-B1 réduisent significativement la liaison aux HDL (lipoprotéines de haute densité).
Un faisceau analogue à trois hélices a été identifié comme site principal de liaison du ligand dans CD36 et LIMP-2.
c. De nombreux ponts disulfure intramoléculaires maintiennent probablement SR-B1 dans une conformation qui prend en charge le transport des HDL, i.e. C280/C334 et C321/C323.
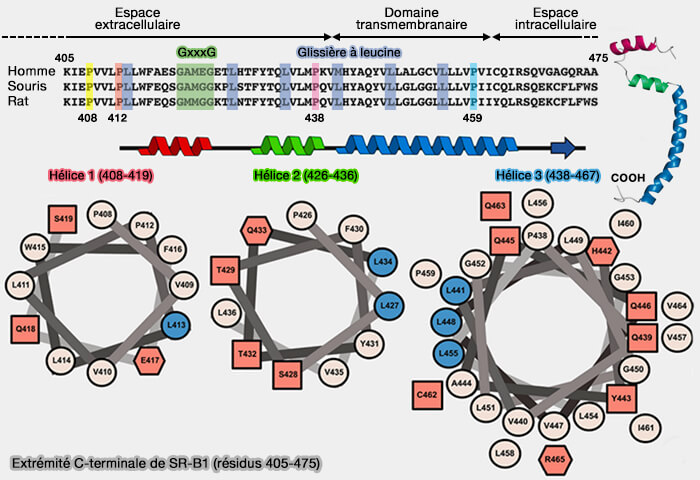
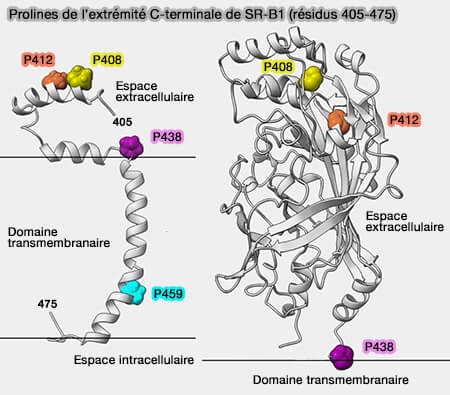
Extrémité C-terminale de SR-B1 (résidus 405-475)
(Figure : vetopsy.fr d'après Chadwick et coll)
P438 pourrait servir de charnière pour une flexibilité accrue de la boucle extracellulaire,
Ces régions extracellulaires assuraient la coordination entre de nombreuses zones de SR-B1 et orientent le récepteur pour une liaison efficace du ligand et le transport du HDL-C, i.e. des mutations perturbent le fonctionnement de SR-B1.
La glycosylation de N108 et N173 est nécessaire pour l'expression de surface de SR-B1.
Domaines transmembranaires (TMD)
Deux domaines transmembranaires α-hélicoïdaux, qui ancrent le domaine extracellulaire, sont courts :
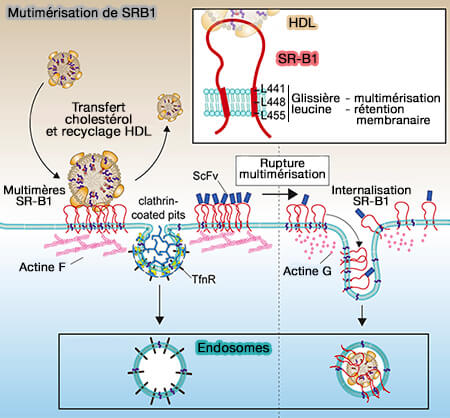
Mutimérisation de SR-B1
(Figure : vetopsy.fr d'après Marques et coll)
le domaine transmembranaire N-terminal (N-TDM) comprend les résidus 13 à 25,
le domaine transmembranaire C-terminal (C-TDM) comprend les résidus 440 à 462.
1. Des domaines de dimérisations, i.e. GxxxG, sont retrouvés dans les deux TDM, mais il semble que seul celui de N-TDM (G12-G15-G18-G25) est susceptible de l'initier, l'autre étant G420-G425.
Des mutations dans cette région ne permettent plus de former des oligomères d'ordre supérieur, corrélées à l'incapacité de se lier aux HDL, d'assure l'absorption des HDL-CE et l'efflux de cholestérol.
L'homo-oligomérisation du SR-B1 est entraînée par des interactions entre les C-TMD.
1. Les extrémités N et C de SR-B1 sont intracytoplasmiques.
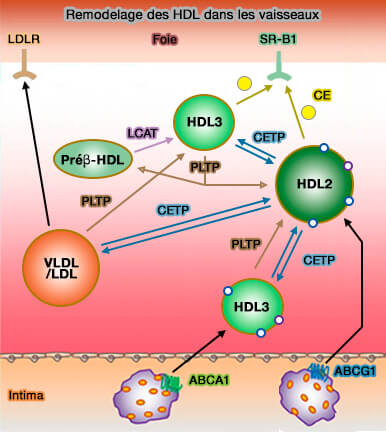
Remodelage des HDL dans les vaisseaux
(Figure : vetopsy.fr d'après Oulmet et coll)
L'extrémité N-terminale, i.e. résidus 1-12, est courte et n'a pas d'utilité précise.
L'extrémité C-terminale, i.e. résidus 463–509, est plus longue.