


Neurophysiologie : synapse
Vésicules synaptiques : cycle vésiculaire
Recyclage vésiculaire
: 6. Endocytose
Processus engagés
- Neurophysiologie
- Cellules gliales et névroglie
- Neurones
- Potentiels membranaires
- Circuits neuronaux
- Neurotransmetteurs
- Récepteurs membranaires
- Synapses
{kind=link}

Dans l'endocytose des vésicules synaptiques, les forces de traction et de constriction de l'actine et de la dynamine, et la séparation de la phase liquide-liquide (LLPS) contribuent à la transition Plat ➞ Λ ➞ Ω ➞ Ο.
Deux processus sont impliqués dans la plupart des modes endocytaires dits dynamine/actine-dépendants pour contribuer à la transition Plat ➞ Λ ➞ Ω ➞ Ο :
- les forces de traction et de constriction de l'actine et de la dynamine,
- la séparation de la phase liquide-liquide (LLPS).
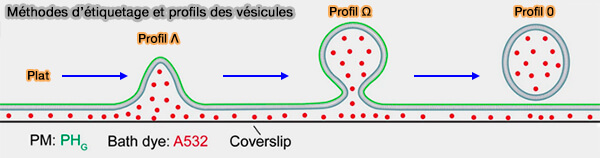
(Figure : vetopsy.fr d'après Wu et Chan)
Traction et constriction
Vue d'ensemble
1. L'actine et la dynamine sont impliquées dans la plupart des modes endocytaires dits dynamine/actine-dépendants pour contribuer à la transition Plat ➞ Λ ➞ Ω ➞ Ο, comme :
- l'endocytose clathrine-dépendante (CME ou clathrin-mediated endocytosis),
- l'endocytose lente,
- l'endocytose ultrarapide (UFE)
- L'endocytose de masse dépendante de l'activité (ADBE ou activity-dependent bulk endocytosis),
- l'endocytose excessive (endocytose > exocytose), overshoot endocytosis,
- l'endocytose indépendante de la clathrine des ligands extracellulaires, des récepteurs, des virus, des bactéries, des prions et des toxines bactériennes (Mechanisms of carrier formation during Clathrin-independent endocytosis 2018).
2. Même pour l'endocytose clathrine-dépendante (CME), la dynamine et l'actine peuvent exercer leurs forces de traction et de constriction pour générer des puits enrobés de clathrine.
- La dynamine pourrait être pré-recruter dans des zones endocytaires par interaction avec la syndapine 1 (
 dynamine et UFE).
dynamine et UFE). - La dynamine est située à la périphérie des patchs de clathrine plats ou peu profonds, avec les epsines, dont EPS15, et FCHO2 qui interviennent dans la courbure de la membrane, ce qui est cohérent avec la capacité de la dynamine à resserrer la base du profil Λ (Endocytic proteins are partitioned at the edge of the clathrin lattice in mammalian cells 2017).
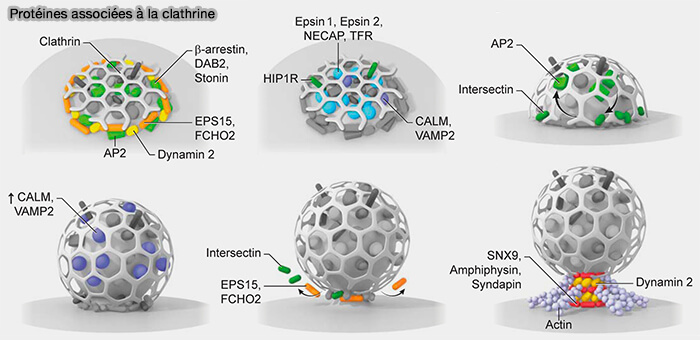
(Figure : vetopsy.fr d'après Sochaki et coll)

La traction et la constriction dépendantes de l'actine et de la dynamine sous-tendent la transformation du profil Plat ➞ Ο, indépendamment de l'implication de protéines de revêtement comme la clathrine.
4. Des influx de calcium variables de faibles à élevés sont impliqués dans la transition Plat ➞ Λ, Λ ➞ Ω et Ω ➞ Ο et augmentent :
a. la vitesse de fermeture des pores Ω-préformés et de fusion, générant une endocytose (Preformed Ω-profile closure and kiss-and-run mediate endocytosis and diverse endocytic modes in neuroendocrine chromaffin cells 2018) :
- lente (plus de ~ 6 s),
- rapide (moins de ~ 6 s),
- ultrarapide (0,6 s),
b. la probabilité de fermeture des pores Ω-préformés et de fusion, générant une endocytose :
- compensatoire (endocytose = exocytose), i.e. compensatory endocytosis,
- excessive (endocytose > exocytose), overshoot endocytosis,
- l'endocytose de masse dépendante de l'activité (ADBE ou activity-dependent bulk endocytosis), qui génère de grandes structures de type endosome, i.e. 100 à 500 nm,
- l'endocytose clathrine-dépendante (CME ou clathrin-mediated endocytosis), formant de petites vésicules, i.e. 40 à 60 nm.
Remarque : plusieurs de ces modes sont également observés dans des cellules non excitables.
(Figure : vetopsy.fr d'après Boltz et coll)
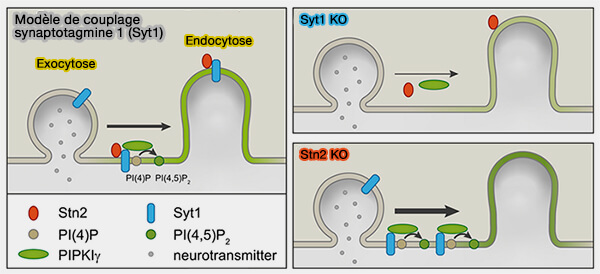
c. La calmoduline, la calcineurine et la protéine kinase C ont été suggérées comme capteurs calciques sous-jacents à l'endocytose déclenchée par la déphosphorylation et/ou la phosphorylation des protéines endocytaires (The Role of Calcium/Calmodulin-Activated Calcineurin in Rapid and Slow Endocytosis at Central Synapses 2010 et Calcineurin Is Universally Involved in Vesicle Endocytosis at Neuronal and Nonneuronal Secretory Cells 2014).
La synaptotagmine 1, un capteur de calcium d'exocytose, peut également être impliqué dans le couplage exo-endocytose ( Syt1 et couplage exocytose/endocytose).
Rôle de l'actine
Au cours de la transition Plat ➞ Λ , la hauteur de Λ augmente avec une protrusion en pic à son sommet tandis que sa base reste constante et est entourée de l'échafaudage d'actine et de dynamine (Preformed Ω-profile closure and kiss-and-run mediate endocytosis and diverse endocytic modes in neuroendocrine chromaffin cells 2021 et Molecular mechanics underlying flat-to-round membrane budding in live secretory cells 2022).
1. La pointe de Λ et sa protubérance en forme de pic sont associées à la croissance de filaments d'actine et de points de dynamine, leurs blocages inhibent la transition Plat ➞ Λ.
L'actine et la dynamine s'enfoncent au centre de la zone endocytaire et à la pointe de l'extrémité de la zone Λ conduisant à la transition Plat ➞ Λ.
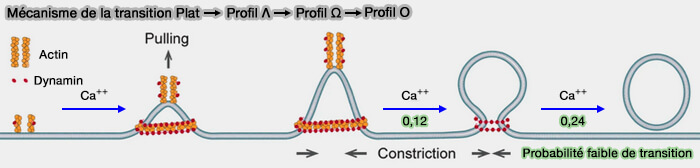
(Figure : vetopsy.fr d'après Shin et coll)
2. Pendant la transition Λ ➞ Ω ➞ Ο, le profil Λ est converti en profil Ω par le rétrécissement de la base.
La dynamine entoure et restreint la base du profil Λ sur des centaines de nanomètres :
- transformant le profil de Λ en Ω, puis
- contractant le pore du profil Ω, i.e. de centaines de nm à 0, en convertissant les profils de Ω en vésicules.
Deux forces, une force de traction au centre et une force de constriction de périphérie à centre, soutenues par l'actine et la dynamine, sont suffisantes pour servir de médiateur à la transition Plat ➞ Λ ➞ Ω ➞ Ο.
3. L'actine, attachée à la pointe du profil Λ en croissance, peut étirer Λ et réduire le pore.
- La modélisation suggère qu'une force d'impulsion ponctuelle de -3 pN est suffisante pour tirer la membrane vers l'intérieur (Molecular mechanics underlying flat-to-round membrane budding in live secretory cells 2022).
- Une telle force pourrait impliquer un faisceau de complexes contractiles d'actomyosine avec un ancrage dans le réseau d'actine cytoplasmique (Actin, a Central Player in Cell Shape and Movement 2009).
Remarque : ce processus diffère de l'endocytose clathrine-dépendante (CME), dans laquelle la polymérisation de l'actine, liée à la cage de clathrine par Hip1 ou Hip1R ou les epsines 1/2/3 par exemple, génère des forces pour allonger le bourgeon ( recrutement de l'actine dans le CME).
Ces mécanismes peuvent expliquer pourquoi l'inhibition de l'actine F par la latrunculine A ou l'activation de l’actine β réduit l'endocytose et les fosses membranaires au niveau des terminaisons nerveuses et inhibe l'endocytose médiée par la clathrine (Actin Is Crucial for All Kinetically Distinguishable Forms of Endocytosis at Synapses 2016 et Synaptic Vesicle Endocytosis Occurs on Multiple Timescales and Is Mediated by Formin-Dependent Actin Assembly 2017).
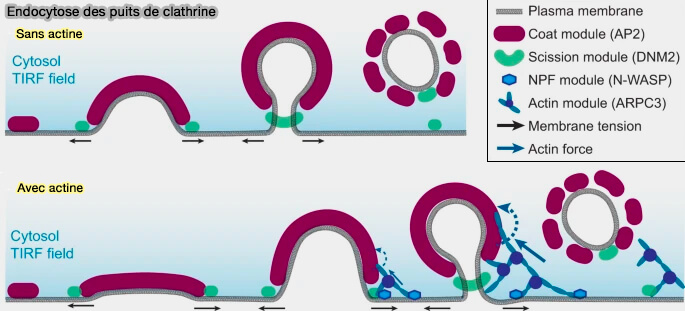
(Figure : vetopsy.fr d'après Jin et coll)
Rôle essentiel de la dynamine
La fission des pores dynamine-dépendante, en rnviron~ 5-20 nm, est bien connue et est étudiée dans un chapitre spécial.
1. Cependant, la dynamine est aussi utilisée dans( traction et constriction) :
- dans la traction de la membrane vers l'intérieur de la vésicule en formation, avec l'actine,
- dans le rétrécissement de la base du profil Λ et du pore.
Remarque : la dynamine et les autres DSP (Dynamin Superfamily Proteins) interviennent dans d'autres fonctions conjointement au cytosquelette.
2. La dynamine seule peut entourer et contracter la base de la vésicule en formant des hélices ( modèles de fission).
Les modèles actuels où les changements de conformation par hélice de la dynamine peuvent se rétrécir de 5-10 nm le pore semblent difficiles à expliquer la constriction de centaines de nanomètres (Structural Insights into the Mechanism of Dynamin Superfamily Proteins 2019).
3. D'autres molécules pourraient également être impliquées, comme la synaptojanine 1 (Synj) ou l'endophiline A ( mécanisme de l'UFE).
Endocytose et LLPS
La séparation de la phase liquide-liquide (LLPS) pourrait contribuer à la génération de courbures endocytaires (Liquid-like protein interactions catalyze assembly of endocytic vesicles 2021).
a. Les protéines de courbures Eps15 et Fcho1/2 reposent sur des interactions faibles et de type liquide pour favoriser l'assemblage de gouttelettes de protéines in vitro, ce qui peut soutenir l'endocytose clathrine-dépendante.
De même, Ede1, l'homologue Eps15 de la levure, peut générer une phase liquide séparée pour nucléer les patchs endocytaires (Condensation of Ede1 promotes the initiation of endocytosis 2022).
b. L'endophiline subit une transition de phase de type liquide en condensats pour faciliter l'assemblage de protéines endocytaires lors de l'endocytose rapide (Multivalent interactions between molecular components involved in fast endophilin mediated endocytosis drive protein phase separation 2022).
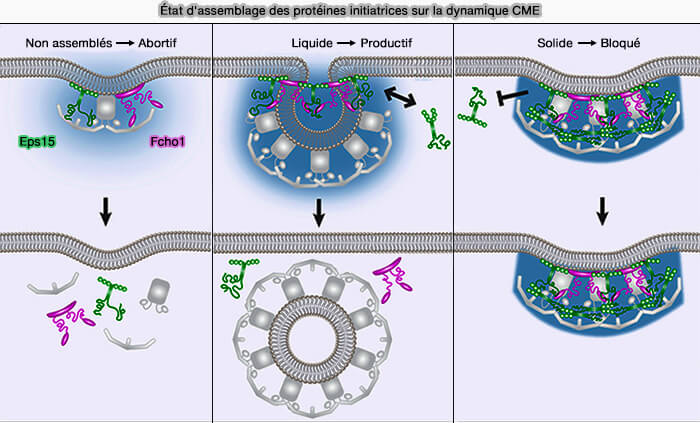
(Figure : vetopsy.fr d'après Day et coll)
2. La dynamine 1 interagit avec la syndapine 1 pour former des condensats moléculaires pour la médiation de l'endocytose ultrarapide (UFE) au niveau des synapses ( dynamine et UFE).
Remarque : les protéines endocytaires de revêtement de domaines de type prions dans les levures peuvent former des puncta hémisphériques avec les caractéristiques des condensats biomoléculaires (Endocytic proteins with prion-like domains form viscoelastic condensates that enable membrane remodeling 2021).
Les interactions cohésives au sein des condensats et les tensions interfaciales entre les condensats, les membranes et le cytosol pourraient contribuer à l'invagination membranaire au cours de l'endocytose dépendante et indépendante de l'actine.
Retour à l'endocytose
NeurophysiologeCellules gliales et névroglieNeuronesPotentiels membranairesCircuits neuronauxNeurotransmetteursRécepteurs membranairesSynapsesStructure synaptiqueNeurotransmissionVésicules synaptiques (VS)Pools synaptiquesRôles du calcium dans l'électrosécrétionCycle synaptiqueTranslocation et attache (tethering)Amarrage (docking) et amorçage (priming)Fusion et exocytoseRecyclage des VSDifférents types d'endocytoseAcidification et remplissage des VSRegroupement des VSPlasticité et régulation synaptique