Vue d'ensemble de la régulation de SREBP par le cholestérol
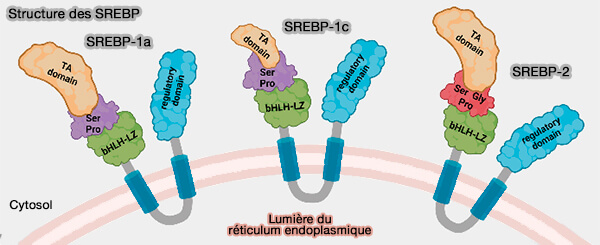
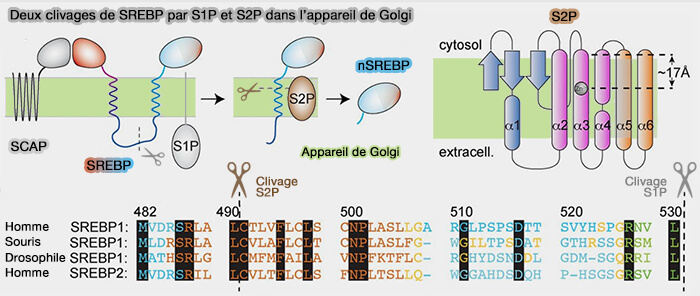
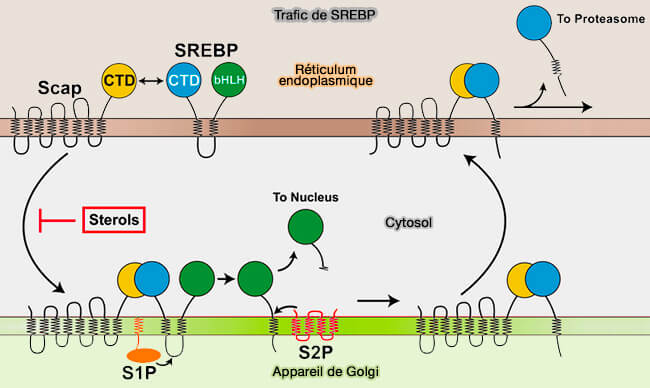
1. Le précurseur SREBP, ancré à la membrane du réticulum endoplasmique (RE), doivent subir deux clivages, i.e. par la protéase du site-1 (S1P) et la protéase du site-2 (S2P) situées dans la membrane de l'appareil de Golgi, pour produire un domaine N-terminal actif, appelé nSREBP ou forme nucléaire de SREBP.
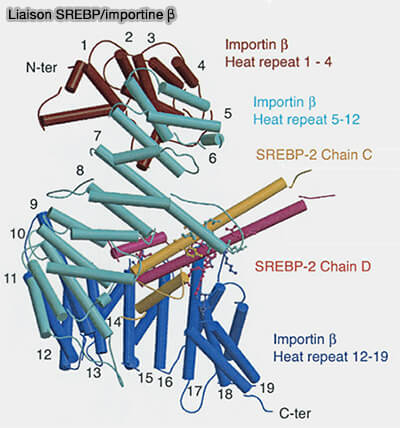
Le domaine bHLH-LZ possède un nouveau signal de localisation nucléaire qui se lie directement à l'importine et permet aux nSREBP d'entrer dans le noyau.
Une fois la translocation dans le noyau, les nSREBP stimulent l'expression de plusieurs gènes associés à la cholestérogénèse et à la lipogénèse.
Structure des SREBP
(Figure : vetopsy.fr d'après Ang et col)
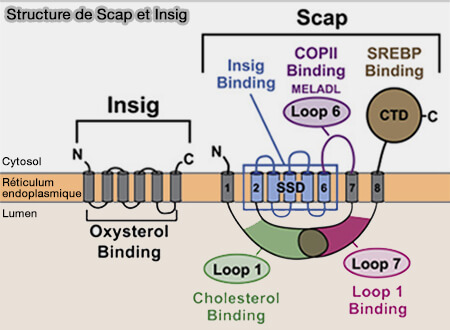
2. La translocation du RE au Golgi implique une autre protéine membranaire du RE appelée SCAP (SREBP cleavage-activating protein).
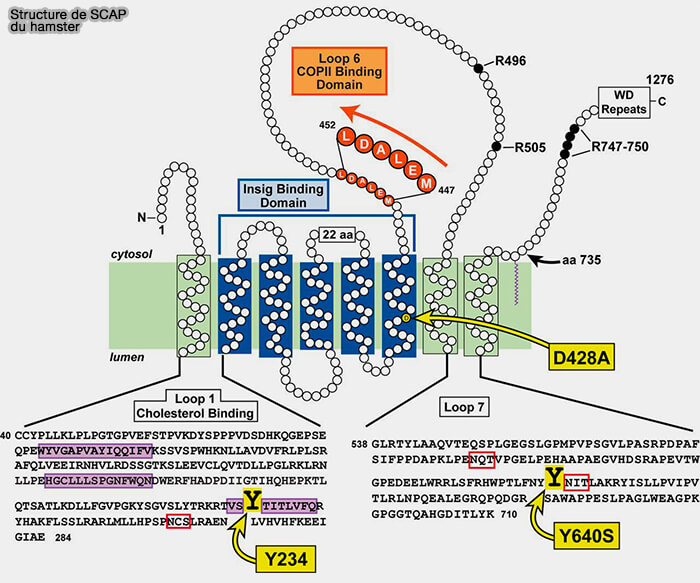
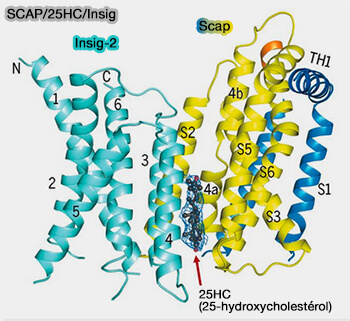
1. La boucle 1 (L1, 240 résidus), localisée dans la lumière du RE, se lie au cholestérol par trois séquences hydrophobes.
Structure de SCAP du Hamster
(Figure : vetopsy.fr d'après Zhang et col)
2. La boucle 7 (L7, 175 résidus environ), localisée dans la lumière du RE, interagit avec la boucle 1, et exerce un rôle régulateur pour le contrôle de la liaison COPII par la boucle L6.
La structure de TM4 (S4), contrairement aux autres TM, est formé par deux demi-hélices, S4a et S4b.
Par rapport aux structures d'autres protéines contenant des SSD comme NPC1 et patched 1, dans lesquelles S4 est droit, l'inclinaison de S4a vers l'intérieur du SSD crée l'espace pour l'hébergement du cholestérol.
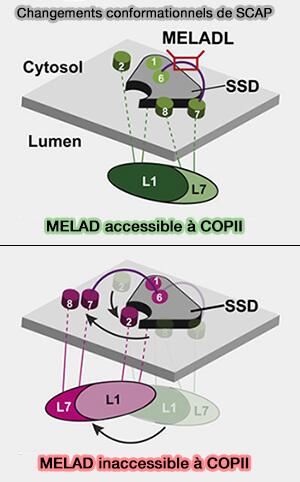
2. Le domaine L1-L7 s'éloigne du module SSD et déplace TM2 et le module TM7-TM8.
Régulation de SREBP
(Figure : vetopsy.fr d'après Kober et coll)
Ce changement conformationnel :
crée une interface de liaison pour Insig-1 ou Insig-2 (Insuline-inducted gene) sur TM2 à TM6.
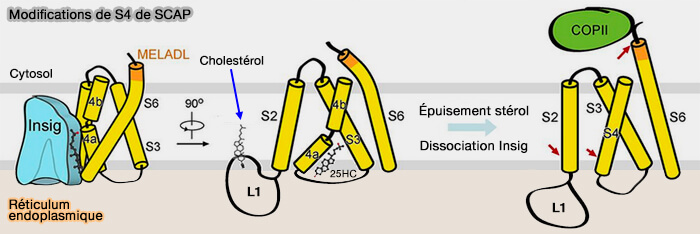
provoque un changement de position de de L6 contenant MELADL du côté cytoplasmique.
3. La formation du complexe ternaire SREBP/SCAP/Insig dans la membrane du RE empêche les SCAP de se lier à COPII et au complexe d'atteindre l'appareil de Golgi pour le clivage de SREBP, supprimant ainsi la transcription des gènes nécessaires à la synthèse du lipides et des LDLR.
Les cellules sont incapables de générer et d'absorber du cholestérol.
pour le déplacement de S2, qui constitue une interface majeure avec Insig.
a. Le 25HC est donc en sandwich entre SCAP (S4, S5 et S6) et Insig (TM2 et TM4) et sa présence stabilise la conformation déroulée de S4 de Scap qui est cruciale pour l'association Insig.
Régulation de SREBP
(Figure : vetopsy.fr d'après Yan et col)
Des mutations dans les Insig qui abolissent la liaison à 25-HC empêchent également la protéine de se lier à Scap et favorisent sa rétention dans les membranes du RE.
Les mutations Scap, telles que D428A (Asp428 → Ala) et Q432A (Gln432 → Ala), qui abaisse le seuil énergétique du déroulement de S4, permettent la formation de complexes avec Insig en l'absence de stérols.
L'interdépendance du déroulement de S4, de l'hébergement 25HC et de la liaison Insig établit la base moléculaire de la détection des stérols.
b. Outre S4, les principaux changements conformationnels de SSD se produisent dans S2 qui est directement connecté à S1.
3. Un modèle simplifié pour l'interaction régulée par le 25HC (25-hydroxycholestérol) entre Scap et Insig pourrait être le suivant.
c. En présence de stérols et Insig, S4 de Scap se déroule, et le motif MELADL est dans une conformation qui ne peut pas être reconnue par COPII.
d. En l'absence des stérols, S4 peut exister sous forme d'hélice intacte comme dans d'autres protéines contenant du SSD. Les changements conformationnels de plusieurs segments SSD exposent le motif MELADL pour la reconnaissance COPII.
Concentration faible de cholestérol
Lorsque de l'épuisement des stérols intracellulaire, SCAP se dissocie des Insigs.
Transport de SREBP vers l'appareil de Golgi
Le domaine globulaire de SCAP formé par L1 et L7 est positionné directement sous le module SSD et le module TM7-TM8 est lié au SSD.
1. Dans cette conformation, la séquence MELADL sur la boucle cytoplasmique L6 est accessible aux protéines COPII.