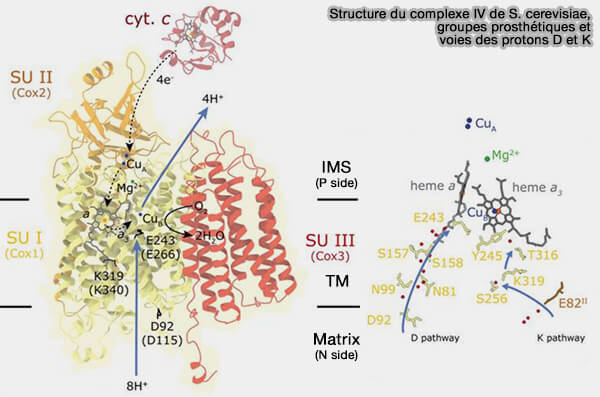
Le complexe IV ou cytochrome c oxydase (CcO) contient trois sous-unités dites catalytiques (I, II et III), contenant des groupes prosthétiques métalliques (Fe et Cu) et un site actif appelé centre binucléaire (BNC).
Structure du complexe IV de S. cerevisiae
(Figure : vetopsy.fr d'après Brzezinski et coll)
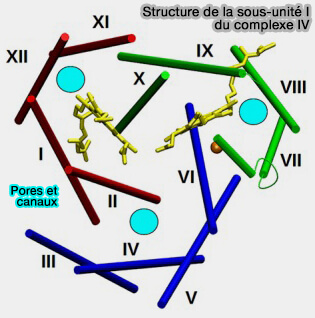
Structure de la sous-unité I
(Figure : vetopsy.fr d'après Wikström et coll)
Le complexe IV (CcO) contient trois sous-unités dites catalytiques.
Sous-unité catalytique I
La sous-unité catalytique I présente un pli hautement conservé.
1. Les 12 hélices transmembranaires sont légèrement inclinées par rapport à la membrane normale et sont disposées en trois groupes avec une quasi-symétrie de type trois.
2. Chacun des trois groupes d'hélices abrite un pore hydrophile, qui contient des résidus d'acides aminés polaires et des molécules d'eau.
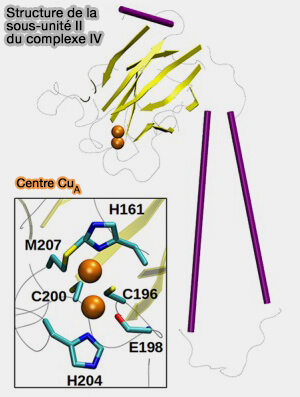
Structure de la sous-unité II
(Figure : vetopsy.fr d'après Wikström et coll)
1. La sous-unité II joue un rôle :
dans le transfert d’électrons,
dans l’absorption des protons par le canal K, grâce au résidu acide conservé, i.e. Glu62 ou 82 dans la deuxième hélice TM.
2. L’interface sous-unité I/II est en partie en liaison avec un ion magnésium (Mg++) et un grand cluster d’eau qui servent de piège transitoire à protons lors de leur pompage.
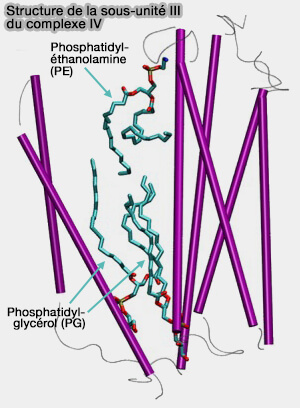
Sous-unité catalytique III
La sous-unité III, hautement conservée, est exclusivement trouvée dans les oxydases de type A.
Elle se compose de 7 hélices TM divisées en deux groupes de 2 et 5 hélices formant chacune une structure en forme de V.
Elle établit des contacts hydrophobes étendus avec la sous-unité I.
La sous-unité III ne contient pas de cofacteurs redox et ne joue pas directement un rôle dans le pompage des protons.
La suppression ou la mutation de la sous-unité III provoque une baisse drastique de l’activité catalytique du complexe IV et même sa désintégration partielle.
1. Elle permet une stabilisation de la structure du complexe IV au sein de la membrane mitochondriale et est nécessaire à l'assemblage et au maintien de la conformation correcte des sous-unités catalytiques (I et II).
Structure de la sous-unité III
(Figure : vetopsy.fr d'après Wikström et coll)
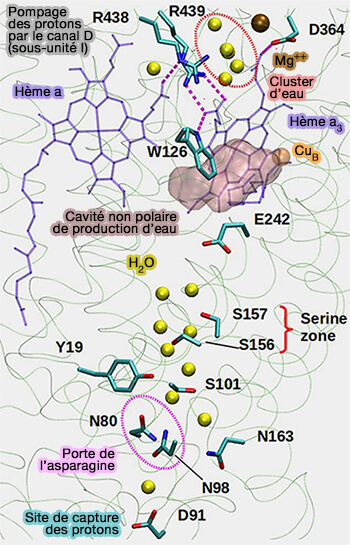
L'inactivation de la CcO est renforcée dans des conditions où l'activité protonique locale près du centre binucléaire (BNC) est faible.
Dans ces conditions, CuB peut acquérir deux hydroxydes (-OH2) et laisser la structure sous forme d'hydroxyde de cuivre.
Un des rôles de la sous-unité III est, pour prévenir cet effet, de permettre une absorption rapide des protons du BNC par le canal D.
a. La région du site de liaison aux lipides jouerait un rôle important dans la canalisation de l’O2 ( voie de l'oxygène et de l'eau).
b. Les groupes de tête chargés des lipides étroitement liés peuvent augmenter le taux de transfert de protons en perturbant le pKa d’Asp91, le site d’absorption des protons du canal D.
Ainsi, elle permettrait une structuration correcte des canaux de pompage des protons.
Structure du site actif : BNC
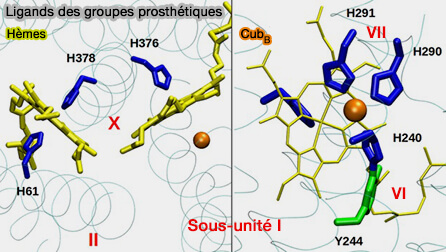
La sous-unité catalytique I de la CcO abrite le site actif de réduction de l'oxygène, i.e. le centre binucléaire (BNC) qui est caractérisé par :
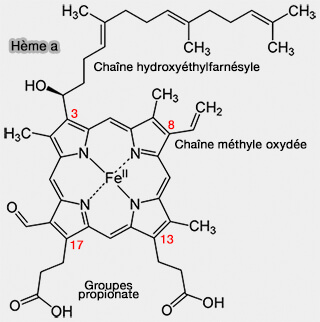
Sur sa face proximale, l'hème fer est lié à une histidine, i.e. H376 hautement conservée de l'hélice X.
Du côté distal, entre le fer et le CuB, on trouve de petits ligands diatomiques tels que CO, CN- et le substrat physiologique O2 qui se lie axialement au Fe.
CuB, l'autre constituant du BNC, est proche de l'hème a3, Fe et Cu ne sont séparés que d'environ 5 Å.
1. L’ion cuivre est lié par :
deux histidines, i.e. H290 et H291 de l’hélice VII de la sous-unité I,
par une autre histidine, i.e. H240 qui est liée de manière covalente à une tyrosine hautement fonctionnelle, i.e. Y244 qui fait partie du ligand unique his-tyr de CuB.
3. L’hème a3 et le CuB interagissent au moyen d’une liaison hydrogène entre l’hydroxyle de la chaîne hydroxyéthylfarnésyle de l’hème a3 et la chaîne latérale de Y244.
Le rôle fonctionnel de cette interaction n’est pas connu, mais il peut être important dans la modulation du pKa de la tyrosine, dont le rôle dans la catalyse est établi ( mécanisme du complexe IV).





 canaux de transfert des protons).
canaux de transfert des protons).