Les synucléines, principalement exprimées au niveau des terminaisons présynaptiques, régulent le trafic de vésicules synaptiques et la libération ultérieure des neurotransmetteurs, mais aussi, jouent un rôle majeur dans les maladies neurodégénératives.
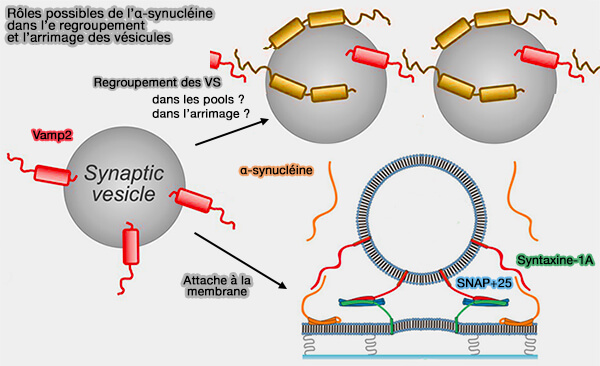
1. Au cours de la neurotransmission, l'α-synucléine (α-Syn) est impliquée dans le regroupement des vésicules synaptiques, entraînant une augmentation locale des vésicules.
6. Les oligomères d'α-syn entraînent un défaut de la fission des vésicules, tandis que les monomères α-syn OE induisent un défaut dans le mécanisme de non-revêtement pendant la CME.
Interactions avec la machinerie
La fusion de la membrane dépendante du complexe SNARE est essentielle pour la libération de neurotransmetteur au niveau de la synapse.
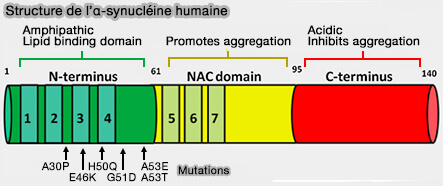
Structure de l'α-synucléine
(Figure : vetopsy.fr d'après Marvin 101)
Remarque : pour ce qui est du regroupement de VS, l'excès d'α-synucléine perturbe la cinétique de formation des condensats synapsine/VS, ce qui indique que le rapport molaire entre la synapsine et l'α-synucléine est important dans leur assemblage (interaction synapsine/α-synucléine).
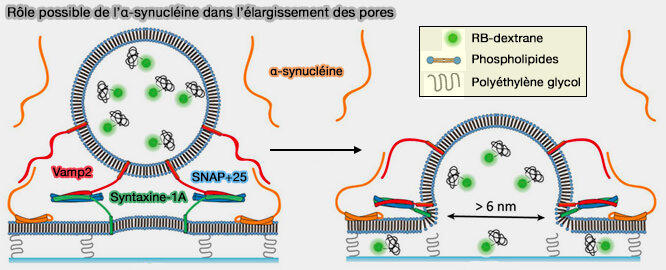
a. Lors de la troncature de l'extrémité C-terminale qui interagit avec le complexe SNARE, le mutant se comporte de la même manière que le type sauvage.
Cependant, les mutants à double proline compromettant la liaison à la membrane montrent des effets significativement réduits sur l'expansion des pores de fusion.
b. L'α-synucléine stimule l'expansion des pores de fusion spécifiquement par sa liaison membranaire.
Rôle possible de l’α-synucléine dans l'élargissement des pores de fusion
(Figure : vetopsy.fr d'après Khoulo et coll)
Fonctions pathologiques des synucléines
Les mutations de l'α-synucléine sont associées à :
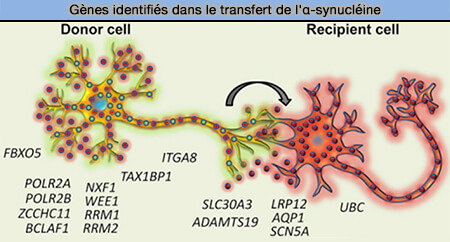
Gènes identifiés dans le transfert de l'α-synucléine
(Figure : vetopsy.fr d'après Kara et coll)
38 gènes régulent le transfert de cellule à cellule de l'α-synucléine, dont ITGA8, un gène candidat identifié par une récente étude d'association à l'échelle du génome PD (GWAS).
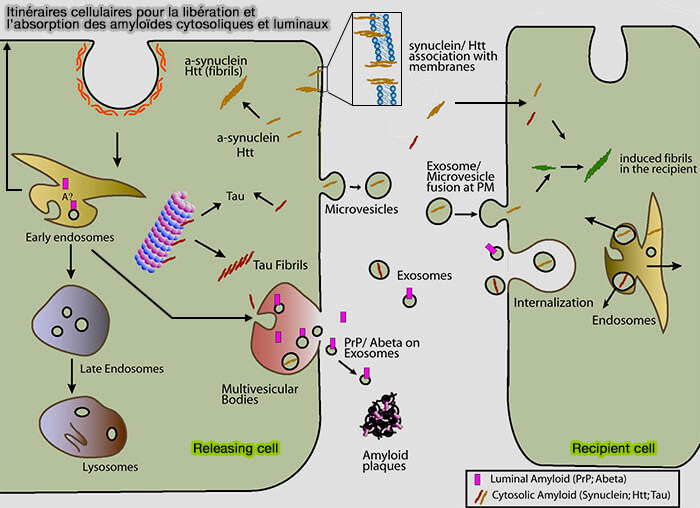
Cette hypothèse montre que des protéines intracellulaires sont non seulement sujettes à l'agrégation pathologique, mais peuvent également être libérées et " infecter " les cellules voisines.
De nombreuses maladies complexes peuvent obéir à un modèle simple de propagation où la pénétration de " grains " ou de " graines ", i.e. seed en anglais, d'ARN ou de protéines dans les hôtes, détermine la propagation spatiale et la progression de la maladie, i.e. ensemencement (seeding model).
Les protéines sont appelées " prionoïdes ", car elles semblent infecter leurs voisins, comme les prions.
Transfert des amyloïdes suivant l'hypothèse prionoïde
(Figure : vetopsy.fr d'après Aguzzi et coll)
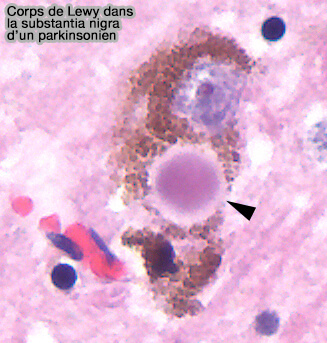
Corps de Lewy
(Figure : vetopsy.fr d'après Tulemo)
Cette maladie emprunte certains symptômes à la maladie d’Alzheimer et à la maladie de Parkinson (Dementia with Lewy bodies).
1. Les corps de Lewy sont des inclusions cytoplasmiques d'α-synucléine mal repliée dans les neurones cérébraux qui sont proches des classiques corps de Lewy retrouvés dans les neurones sous-corticaux de la maladie de Parkinson (Lewy bodies 2006).
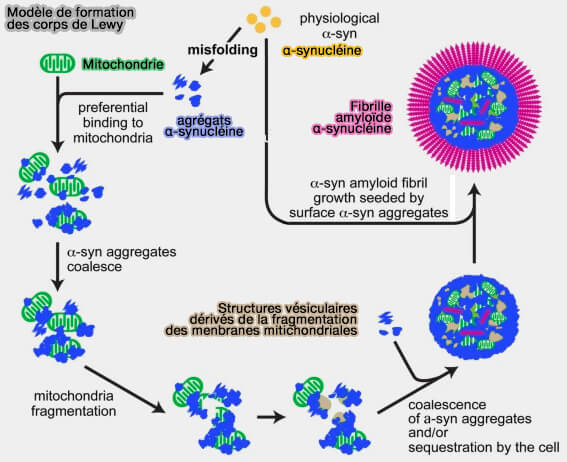
Contrairement aux αSYN physiologiques, les agrégats d'αSYN pathogènes se lient préférentiellement aux mitochondries, indiquant un dysfonctionnement mitochondrial comme mécanisme en aval commun pour les α-synucléinopathies.
Modèle de formation des corps de Lewy
(Figure : vetopsy.fr d'après Wang et coll)



{kind=link}


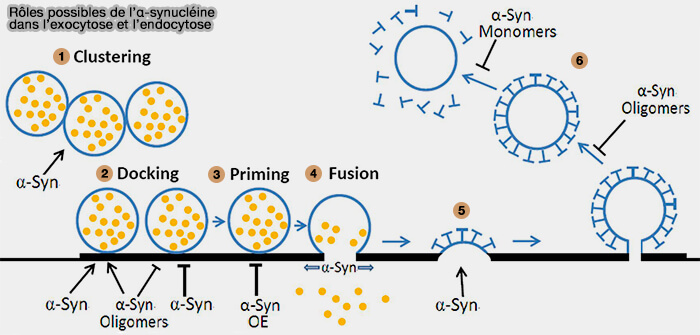
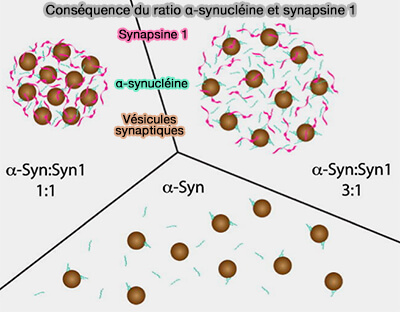
 interaction synapsine/α-synucléine).
interaction synapsine/α-synucléine).