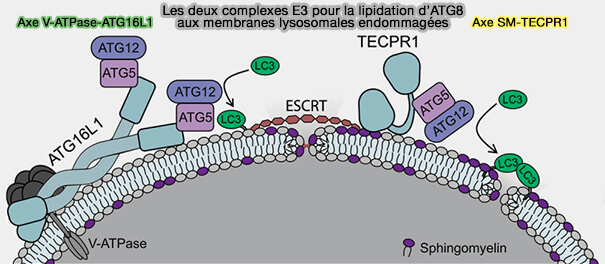
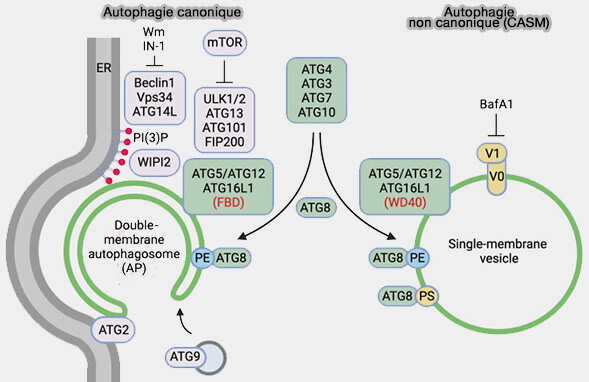
3. Ces deux autophagies impliquent des processus complètement différents, même si elles parviennent toutes deux à la lipidation de LC3.
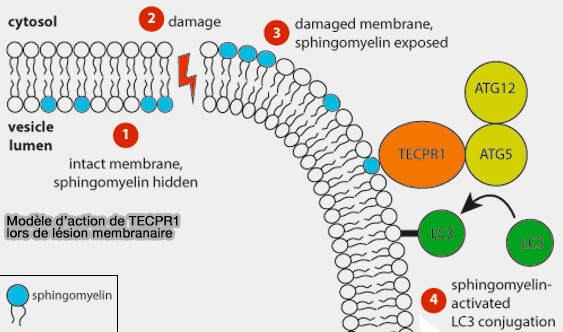
Autophagie non canonique : axe SM-TECPR1
Peut-être dans le but d’augmenter le répertoire d’événements lipidateurs d’ATG8, les vertébrés ont développé deux partenaires complexes alternatifs pour ATG5-ATG12, i.e. :
ATG16L2, qui pourrait entrer en compétition avec ATG16L1 pour la formation de complexe avec ATG12-ATG5 et réduirait l'autophagie,
deux domaines dysferline (DysF) Insérés entre la première et la deuxième pale de chaque hélice, qui semblent reconnaître les membranes sphingomyéline‐positives, i.e. sphingomyéline (SM) qui normalement se trouve sur le feuillet luminal.
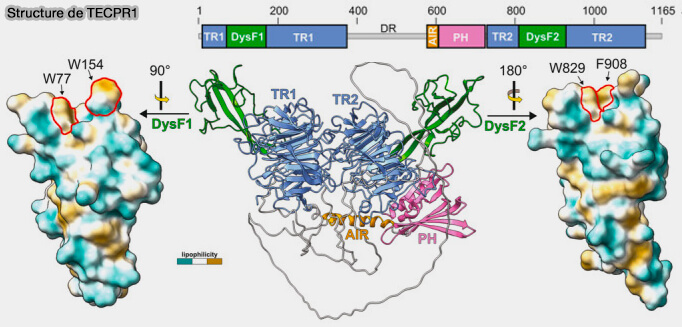
Structure de TECPR1
(Figure : vetopsy.fr d'après Kaur et coll)
L’interaction de LC3C et de TECPR1 favorise le recrutement des autophagosomes vers les lysosomes pour la dégradation lors de l'autophagie sélective.
L’expression accrue de TECPR1 dans les cellules souches neurales réduit le nombre d’agrégats de protéines en favorisant leur clairance autophagique, tandis que l’inhibition de LC3C inhibe l’agrégation.
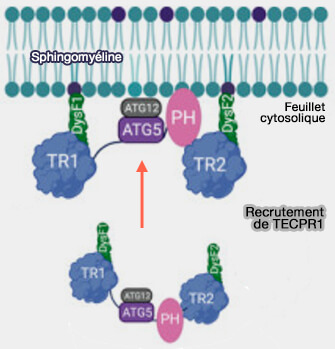
b. Le domaine PH de TECPR1 interagit avec PI(4)P pour cibler TECPR1 vers les lysosomes.
L’échange du domaine PH contre un domaine tandem-FYVE cible TECPR1 aux endosomes et conduit à une accumulation d’autophagosomes LC3C au niveau des endosomes et empêche leur livraison aux lysosomes.
Remarque : TECPR1 pourrait recruter un facteur tardif nécessaire pour la fusion autophagosome-lysosome par son partenaire putatif TRAPPC12 (rôles de TRAPP-III dans l'autophagie).
Recrutement de TEPCR1
(Figure : vetopsy.fr d'après Tan et Finkel)
ATG16L1 le ferait par les résidus de son hélice amphipathique, hélice 2 ( interaction d'ATG16L1).
TECPR1 ne possède pas de structure ou de séquence similaire, mais l'hélice 2 pourrait être située dans le domaine PH de TECPR1.
Ce domaine est plus hydrophobe que la plupart des autres membres de la grande famille des domaines PH, en particulier dans sa région variable de la boucle 1.
un domaine PH qui attache fermement la protéine aux membranes via des interactions hydrophobes.
L’activité de liaison membranaire du domaine PH doit être inhibée en l’absence de sphingomyéline (SM) cytosolique.
Globalement, le rôle des domaines DysF serait alors de reconnaître les SM pour ouvrir la structure protéique afin de permettre l’accès membranaire du domaine PH.
2. Cependant, la réparation des lésions endolysosomales est essentielle pour éviter les fuites d’ions et d’hydrolases dans le cytosol, pour éviter la lysophagie ou pire, l'activation de la mort cellulaire.
leurs domaines PHP qui se lient au PI(4)P lysosomal.
leurs domaines ORD de liaison lipidique de l’ORP1L au cholestérol et de l’OSBP à PI(4)P.
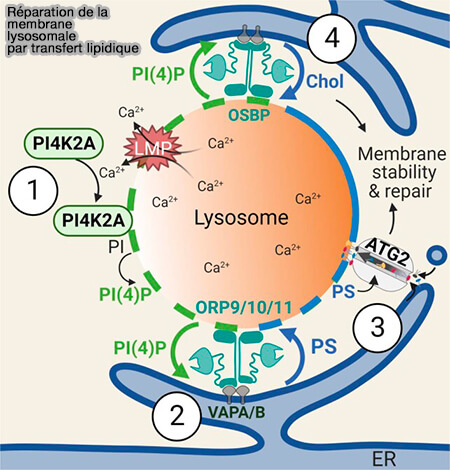
2.4. Le transfert des lipides vers le lysosome favorise la réparation membranaire.
a. ORP1L transfère le cholestérol du RE.
En raison de la capacité intrinsèque du cholestérol à améliorer la rigidité et la stabilité de la membrane, l’accumulation de cholestérol lysosomal pourrait aider directement à la réparation de la membrane.
L’excès de PI(4)P est transporté du lysosome vers le RE par l’OSBP et ensuite déphosphorylé par SAC1.
b. ORP9/10/11 transfère la phosphatidylsérine (PS) qui active Atg2 qui fournit de grandes quantités de lipides (3).




 rôles de TRAPP-III dans l'autophagie).
rôles de TRAPP-III dans l'autophagie).