


Corps nucléaires
1. Corps liés à la biogenèse des ARN
1.2. Corps de Cajal
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Matrice extracellulaire
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires

Le corps de Cajal est un compartiment nucléaire non membranaire impliqué dans la maturation et l’assemblage des snRNP et dans la régulation de la biogenèse des complexes ribonucléoprotéiques (RNP).
Structure et organisation des corps de Cajal
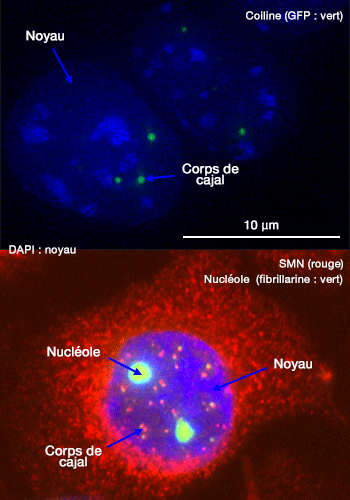
Les corps de Cajal ou corps pelotonnés (CB : Cajal Body ou anciennement " Coiled Bodies ") sont des structures nucléaires sphériques de 0,1 à 2,0 µm de diamètre, présentes à raison de 0 à 10 par noyau (The Cajal body 2008 et The Cajal Body and Histone Locus Body 2010).
(Figure : vetopsy.fr d'après Martin Machyna et
{kind=link}
BioMed Central Open access)
{kind=link}
Les corps de Cajal sont surtout observés dans les cellules à forte activité proliférative (cellules embryonnaires, tumorales) et dans certaines cellules très actives sur le plan métabolique, comme les neurones, où ils ont été décrits près du nucléole et initialement désignés « nucleolar accessory bodies » par Santiago Ramón y Cajal en 1903).
- Les corps de Cajal, comme les histone locus bodies, sont généralement détectables tout au long de l’interphase, puis se désassemblent lors de la mitose.
- Ils ne sont pas strictement indispensables à la viabilité cellulaire dans de nombreuses conditions basales.
- Toutefois, ils optimisent l’efficacité de la maturation et de l’assemblage des complexes ribonucléoprotéiques (RNP) nucléaires et leur contribution devient plus critique lorsque la demande en biogenèse/traitement des RNP augmente ou en contexte de stress.
Composants moléculaires des corps de Cajal
Les corps de Cajal sont constitués d’un ensemble spécifique de protéines structurales, de facteurs d’assemblage des ribonucléoprotéines et d’ARN spécialisés, qui assurent leur formation, leur organisation et leur fonction dans la maturation et le trafic des RNP nucléaires (Cajal’s organization of neuronal nucleus revisited 2025).
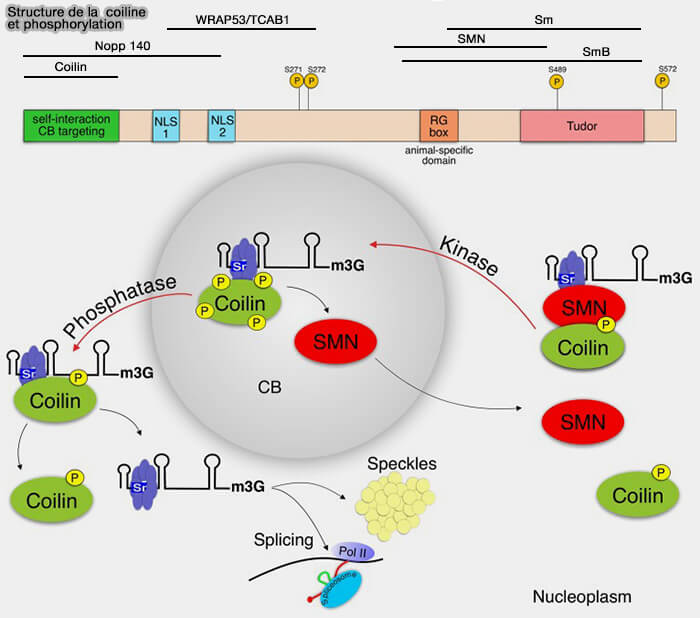
Coilline : protéine structurale centrale
La coiline, protéine structurale centrale et marqueur principal des corps de Cajal, interagit avec de nombreux composants des ribonucléoprotéines nucléaires (RNP) nucléaires et constitue l’échafaudage moléculaire nécessaire à la formation et à l’organisation de ce compartiment.

Vous pouvez lire : Coilin and Cajal bodies (2023).
1. Elle joue un rôle essentiel dans (Cajal-body formation correlates with differential coilin phosphorylation in primary and transformed cell lines 2009) :
- la nucléation des corps de Cajal,
- leur organisation structurale,
- leur dynamique intranucléaire.
(Figure : vetopsy.fr d'après Taliansky et coll)
2. L’état de phosphorylation de la coiline régule sa capacité d’auto-association et conditionne ainsi la formation et la stabilité des corps de Cajal selon le type cellulaire et l’état physiologique.
- Dans les cellules primaires présentant peu ou pas de CB, la coiline est fortement phosphorylée sur ses résidus sérine C-terminaux, ce qui limite son auto-association et la formation de CB.
- Dans les cellules transformées ou fortement prolifératives, la coiline est moins phosphorylée, ce qui favorise son auto-association, la nucléation et le maintien des corps de Cajal.
L’état de phosphorylation de la coilinemodule ses interactions avec le complexe SMN et les snRNP, contribuant à leur recrutement, leur maturation et leur libération.
Autres protéines du corps de cajal
Outre la coiline, les corps de Cajal contiennent de nombreuses protéines impliquées dans l’assemblage, la maturation, le trafic et la modification des complexes ribonucléoprotéiques (RNP) nucléaires.
1. Certaines protéines participent principalement à la biogenèse et à la maturation des snRNP spliceosomaux.
- La protéine SMN (Survival of Motor Neuron), associée aux protéines GEMIN, constitue un complexe multiprotéique essentiel à l’assemblage du cœur Sm des snRNP spliceosomaux.
- Le complexe Integrator participent également à la biogenèse des snARN en assurant le clivage de leur extrémité 3′ co-transcriptionnelle et en contribuant au couplage entre transcription et maturation des snRNP.
2. D’autres protéines interviennent dans l’organisation fonctionnelle des RNP nucléaires et dans les modifications post-transcriptionnelles des snARN.
- WRAP53/TCAB1 (Telomerase Cajal Body Protein 1) reconnaît notamment la boîte CAB présente sur certains scaARN et sur l’ARN télomérase TERC, assurant leur adressage vers les corps de Cajal où se déroulent différentes étapes de maturation et d’assemblage ribonucléoprotéique.
- Nopp140 (Nucleolar phosphoprotein of 140 kDa) agit comme une protéine d’échafaudage hautement phosphorylée capable d’interagir simultanément avec plusieurs composants des scaRNP et des snRNP, favorisant leur concentration dans les corps de Cajal ainsi que l’efficacité des modifications post-transcriptionnelles des snARN.
- La fibrillarine et la dyskérine, protéines respectivement associées aux complexes de type C/D et H/ACA, participent aux principales modifications post-transcriptionnelles des snARN spliceosomaux, 2′-O-méthylation, et pseudouridylation respectivement.
Biogenèse et maturation des snRNP
Les corps de Cajal sont impliqués dans la biogenèse, la maturation et le trafic intranucléaire de plusieurs classes de ribonucléoprotéines nucléaires ou RNP (Cajal bodies: Evolutionarily conserved nuclear biomolecular condensates with properties unique to plants 2023) :
- les snRNP spliceosomaux,
- le snRNP U7 impliqué dans la maturation des ARN d’histones et fonctionnellement associé aux histone locus bodies,
- les scaRNP,
- certaines snoRNP qui peuvent également y transiter au cours de leur maturation.
Le développement suivant concerne principalement la biogenèse et la maturation des snRNP spliceosomaux majeurs, U1, U2, U4 et U5.
- Le snRNP spliceosomal U6 possède une maturation essentiellement nucléaire, sans export cytoplasmique, et s’associe à un anneau de protéines LSm2-8.
- Le snRNP U7 impliqué dans la maturation des pré-ARNm d’histones réplicatifs présente également une biogenèse particulière distincte des snRNP spliceosomales majeures avec une maturation essentiellement nucléaire et contient un cœur Sm atypique dans lequel les protéines LSM10 et LSM11 remplacent SmD1 et SmD2 (
 mécanisme de la maturation des ARNm d’histones réplicatifs).
mécanisme de la maturation des ARNm d’histones réplicatifs).
Transcription et maturation initiale des snARN
Les snARN spliceosomaux majeurs U1, U2, U4 et U5 sont principalement transcrits par l’ARN polymérase II, tandis que le snARN U6 est transcrit par l’ARN polymérase III (Molecular mechanism of quality control during snRNP biogenesis).
1. Les transcrits snARN synthétisés par l’ARN polymérase II acquièrent rapidement une coiffe 5′ monométhylguanosine (m7G), reconnue par le complexe CBC (Cap-Binding Complex).
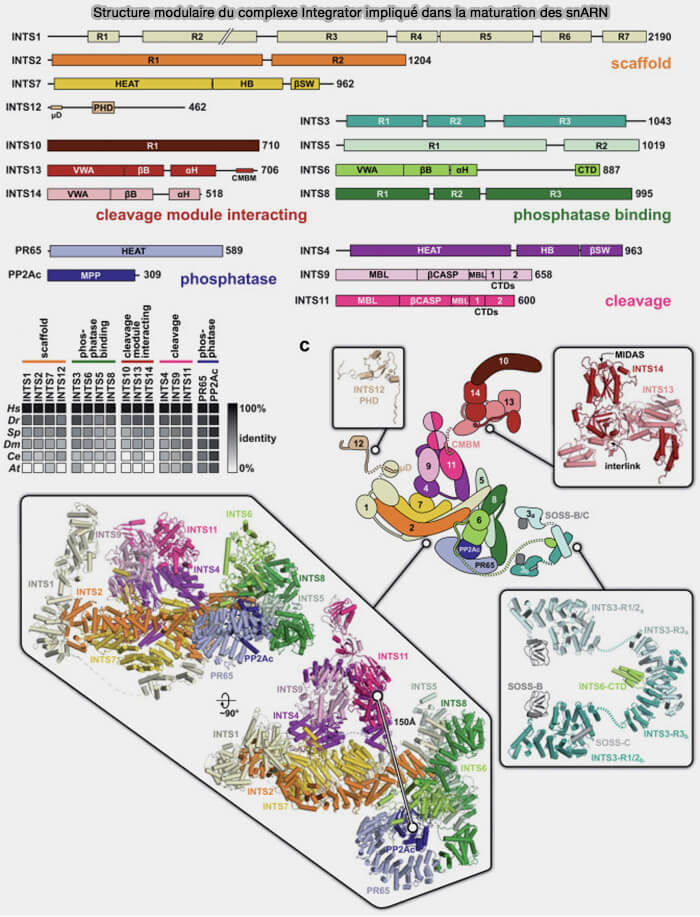
2. Le complexe Integrator est un large complexe multiprotéique constitué d’au moins quatorze sous-unités (INTS1 à INTS14) associées au domaine C-terminal (CTD) phosphorylé de l’ARN polymérase II, où il coordonne la maturation co-transcriptionnelle des snARN (Take a break: Transcription regulation and RNA processing by the Integrator complex 2022 et The integrator complex is required for integrity of Cajal bodies 2012).
(Figure : vetopsy.fr d'après Sabath et Jonas)
a. Le complexe Integrator reconnaît des séquences spécifiques situées en aval des gènes de snARN transcrits par l’ARN polymérase II et recrute une activité endonucléolytique responsable du clivage de l’extrémité 3′ des pré-snARN (Structural basis of the Integrator complex assembly and association with transcription factors 2024).
- Cette activité est principalement associée à la sous-unité catalytique INTS11, une endonucléase de type β-CASP/MBL (CPSF-Artemis-SNM1-PSO2/Metallo-β-lactamase) du module de clivage (INTS4/9/11), apparentée à CPSF73 impliquée dans le clivage des pré-ARNm polyadénylés.
- Contrairement aux ARN messagers, les snARN spliceosomaux ne subissent généralement pas de polyadénylation stable, et leur maturation dépend donc d’un mécanisme spécifique de clivage assuré par Integrator.
b. Certaines sous-unités du complexe Integrator, notamment INTS4, contribuent au recrutement et à la stabilisation du complexe Integrator sur les transcrits snARN associés à l’ARN polymérase II, facilitant le clivage correct de l’extrémité 3′ des pré-snARN.
Une altération de cette maturation perturbe l’accumulation normale des snRNP et entraîne une désorganisation des corps de Cajal, soulignant le lien fonctionnel étroit entre maturation des snARN et intégrité des voies de biogenèse des snRNP nucléaires.
Remarque : outre son rôle dans la maturation des snARN, le complexe Integrator intervient également dans la régulation de la pause transcriptionnelle proximale et de l’atténuation transcriptionnelle de certains gènes transcrits par l’ARN polymérase II (pause proximale de l’ARN polymérase II).
3. Après cette maturation initiale nucléaire, les snARN sont exportés vers le cytoplasme où débute l’assemblage des snRNP spliceosomaux.
- Les snARN ne possèdent pas de signal peptidique d’export nucléaire de type NES (Nuclear Export Signal).
- Leur export dépend donc principalement de PHAX (Phosphorylated Adapter for RNA Export), une protéine adaptatrice qui reconnaît le complexe CBC associé à la coiffe m7G des snARN et recrute l’exportine CRM1/XPO1 en présence de Ran-GTP ( export nucléocytoplasmique).

(Figure : vetopsy.fr d'après Taliansky et coll)
Assemblage cytoplasmique des snRNP
Après leur export nucléaire, les snARN spliceosomaux transcrits par l’ARN polymérase II sont pris en charge dans le cytoplasme par le complexe SMN (Survival of Motor Neuron).
Vous pouvez lire : The SMN complex drives structural changes in human snRNAs to enable snRNP assembly (2023).
1. Le complexe SMN est organisé autour de la protéine SMN, capable de former des oligomères - souvent décrits comme des dimères fonctionnels - associés à plusieurs protéines GEMIN (GEMIN2 à GEMIN8), qui participent à l’assemblage, à la stabilisation et au contrôle qualité des snRNP spliceosomaux (Diverse role of Survival Motor Neuron Protein 2017).
GEMIN2, également appelée SIP1 (SMN-interacting protein 1), joue notamment un rôle central dans l’organisation des protéines Sm autour des snARN, tandis que GEMIN3 à GEMIN7 participent à différentes étapes d’assemblage et de remodelage des complexes ribonucléoprotéiques (Structure of a Key Intermediate of the SMN Complex Reveals Gemin2’s Crucial Function in snRNP Assembly 2012).
2. Dans le cytoplasme, plusieurs protéines SM, SmB/B′, SmD1 et SmD3, sont prises en charge par le complexe méthylosome contenant PRMT5/MEP50 qui catalyse la diméthylation symétrique de certaines arginines, modification favorisant leur reconnaissance par le complexe SMN-GEMIN (The SMN Complex: An Assembly Machine for RNPs 2006).
Des sous-complexes Sm préassemblés comprenant notamment SmD1/D2 et SmE/F/G sont ensuite pris en charge transitoirement par la protéine chaperon pICln avant leur transfert vers la machinerie SMN-GEMIN d’assemblage des snRNP
Les protéines Sm possèdent naturellement une forte tendance à l’oligomérisation spontanée et à l’agrégation et pICln agit ainsi comme un chaperon empêchant l’assemblage prématuré des anneaux Sm.
3. Le complexe SMN-GEMIN agit ensuite comme une plateforme d’assemblage et de contrôle qualité des snRNP spliceosomaux (The SMN complex drives structural changes in human snRNAs to enable snRNP assembly 2023).
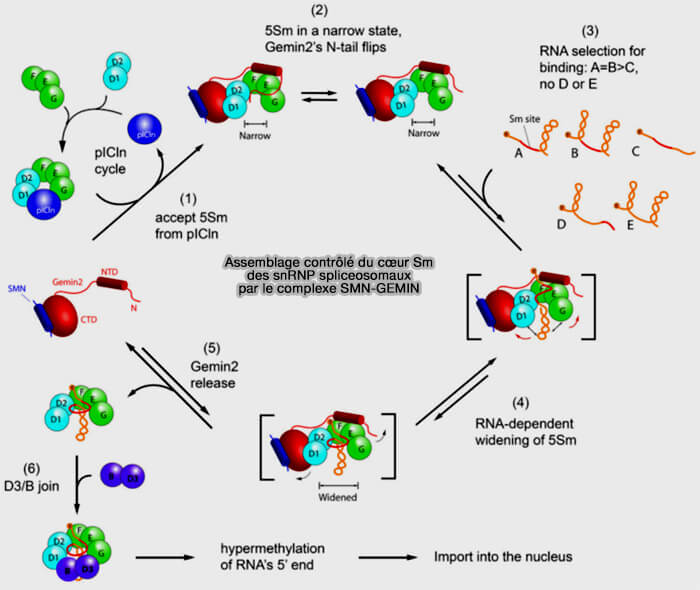
a. Un complexe intermédiaire appelé " 5Sm ", formé par les protéines SmD1, SmD2, SmE, SmF et SmG, est positionné sur le complexe SMN-GEMIN.
GEMIN2 joue ici un rôle central en maintenant ces protéines Sm dans une conformation enfer à cheval, i.e ouverte , empêchant la fermeture prématurée de l’anneau (Structure of a Key Intermediate of the SMN Complex Reveals Gemin2's Crucial Function in snRNP Assembly 2011).
(Figure : vetopsy.fr d'après Yi et coll)
b. GEMIN5 reconnaît ensuite le code snRNP du snARN qui comprend (Understanding GEMIN5 Interactions: From Structural and Functional Insights to Selective Translation 2025) :
- le site Sm riche en uridines, à séquence consensus AUUUUUG et à conformation simple brin,
- une structure secondaire adjacente de type tige-boucle (The SMN Complex: An Assembly Machine for RNPs 2002).
La figure tirée de cet article ancien montre le complexe entier, alirs que c'est le complexe intermédiaire appelé " 5Sm ".
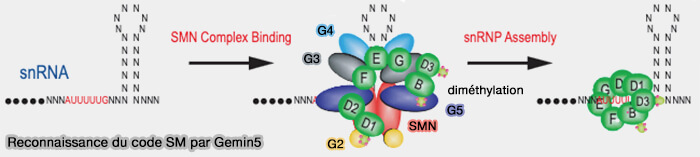
(Figure : vetopsy.fr d'après Battle et coll)
c. Le snARN reconnu par GEMIN5 est ensuite engagé dans ce pré-complexe de protéines Sm maintenu dans une conformation ouverte par GEMIN2.
- Le site Sm s’insère progressivement dans la cavité du pré-complexe Sm et modifie la géométrie, passant d’un état étroit, narrow, à un état élargi, widened, ce diminue l’affinité de GEMIN2 ou negative cooperativity (Negative cooperativity between Gemin2 and RNA provides insights into RNA selection and the SMN complex's release in snRNP assembly 2020).
- L’écartement des protéines SmD1/D2 et SmE/G, permettant l’incorporation finale de SmD3 et SmB/B′ qui ferment l’anneau heptamérique autour du site Sm du snARN.
d. Le cœur Sm mature ainsi stabilisé diminue progressivement l’affinité du complexe SMN-GEMIN pour la particule assemblée, entraînant sa dissociation
4. La formation correcte du cœur Sm constitue un signal de maturation permettant le recrutement de la triméthylguanosine synthase TGS1.
- La coiffe 5′ monométhylguanosine (m7G) des snARN est alors hyperméthylée en coiffe triméthylguanosine (TMG), modification caractéristique des snRNP spliceosomaux.
- TGS1 ajoute deux groupements méthyle supplémentaires sur l’azote N2 de la guanosine coiffante et on passe donc de m7G à m32,2,7G (TMG cap).
Le snARN U6 constitue une exception car, étant transcrit par l’ARN polymérase III, il ne possède pas de coiffe triméthylguanosine (TMG) classique des snRNP spliceosomaux majeurs transcrits par l’ARN polymérase II, mais une coiffe coiffe γ-monométhylphosphate en 5'.
Remarque : la coiffe triméthylguanosine (TMG) n’est pas exclusive aux snRNP spliceosomaux et peut également être retrouvée sur plusieurs autres ARN nucléaires associés à des complexes ribonucléoprotéiques spécialisés, notamment certains snoARN, scaARN et le snARN U7.
5. La formation du cœur Sm et l’hyperméthylation de la coiffe TMG favorisent la reconnaissance des pré-snRNP par le domaine siBB de la snurportine 1 (SPN1/SNUPN), adaptateur d’import nucléaire qui interagit avec la coiffe triméthylguanosine et recrute l’importine β responsable du transport nucléaire des pré-snRNP.
Remarque: la protéine SMN a initialement été observée dans des structures nucléaires distinctes mais étroitement associées aux corps de Cajal, appelées Gems (Gemini of Cajal bodies), d'où leur nom en relation avec les gémaux, qui sont morphologiquement similaires, mais dépourvues de coiline.
Dans la plupart des cellules, SMN et coiline colocalisent dans les corps de Cajal, tandis que les Gems correspondent probablement à des structures dynamiques contenant des complexes SMN engagés dans différentes étapes de la biogenèse et du recyclage des snRNP.
6. Outre le cœur Sm, les snRNP spliceosomaux contiennent également des protéines spécifiques propres à chaque particule (snRNP-specific proteins), généralement nommées selon la particule spliceosomale à laquelle elles sont associées, qui déterminent leurs fonctions au sein du spliceosome.
Certaines de ces protéines commencent à s’associer aux snRNP dès les étapes cytoplasmiques de maturation, tandis que d’autres sont recrutées ou remodelées après le réimport nucléaire dans les corps de Cajal ( protéines des snRNP).
- Par exemple, les protéines spécifiques du snRNP U1 - U1-70K, U1A et U1C - peuvent s’associer précocement au cours de la biogenèse cytoplasmique des snRNP.
- Certaines protéines associées au snRNP U2, comme U2A′ et U2B", sont principalement recrutées lors des étapes nucléaires de maturation dans les corps de Cajal.

(Figure : vetopsy.fr d'après Smoliński et coll)
Maturation des snRNP dans les corps de Cajal : scaARN et scaRNP
Après leur réimport nucléaire, les pré-snRNP transitent par les corps de Cajal, qui constituent des sites spécialisés de maturation, de modification et de contrôle qualité des snARN spliceosomaux.
1. Chez les eucaryotes, les scaRNP (small Cajal body-specific ribonucleoproteins) sont des complexes ribonucléoprotéiques spécialisés proches des snoRNP nucléolaires qui contiennent les scaARN (small Cajal body-specific RNA), une classe spécialisée de petits ARN non codants apparentés aux snoARN (Towards an understanding of regulating Cajal body activity by protein modification 2016).
a. Ces scaARN possèdent des caractéristiques structurales similaires aux snoARN, impliqués dans la maturation des ARNr qui s’accumulent principalement dans le nucléole, incluant les motifs conservés CD et H/ACA.
- Les scaRNP de type C/D contiennent notamment la fibrillarine, qui catalyse la 2′-O-méthylation, NOP56, NOP58 et SNU13/15.5K, protéines impliquées dans l’assemblage, la stabilité structurale et la reconnaissance des scaARN contenant des motifs C/D.
- Les complexes de type H/ACA possèdent principalement la dyskérine, qui catalyse la pseudouridylation, associée à NOP10, NHP2 et GAR1, protéines qui stabilisent les ARN H/ACA et participent à l’organisation structurale et à l’activité fonctionnelle des complexes scaRNP.
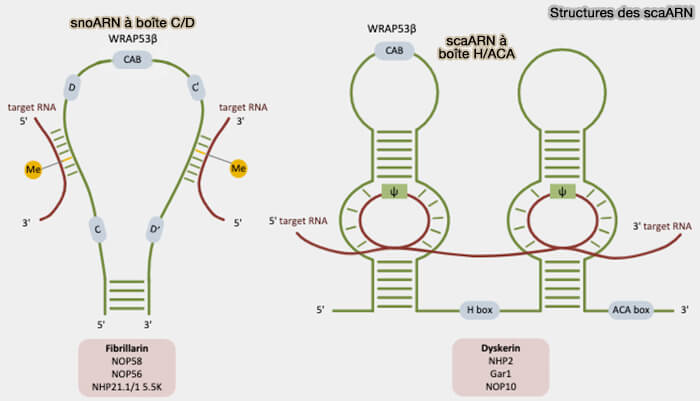
(Figure : vetopsy.fr adaptée d'après Coucoravas)
b. Certaines scaRNP contiennent également des protéines Sm associées à des motifs de type site Sm présents sur certains scaARN, contribuant à leur stabilité et à leur localisation dans les corps de Cajal.
c. Des protéines spécifiques de certaines snRNP, comme U2B" et U2A′ associées au snRNP U2.
Remarque : les corps de Cajal interviennent également dans certaines étapes de la biogenèse et de la maturation des snoRNP, en particulier des complexes contenant des ARN à boîte H/ACA.
(Figure : vetopsy. fr d'après Lamon)
2. Cependant, les scaARN possèdent également une boîte CAB (Cajal body box), reconnue par la protéine WRAP53/TCAB1, qui assure leur ciblage vers les corps de Cajal et guident les principales modifications post-transcriptionnelles des snARN spliceosomaux (WRAP53β and scaRNA2 on their journey to repair the DNA 2018).
3. Les modifications post-transcriptionnelles des snARN spliceosomaux contribuent à :
- la stabilité structurale des snARN,
- leur repliement correct,
- à l’organisation fonctionnelle des futurs complexes spliceosomaux.
4. Après leur maturation finale, les snRNP sont redistribués vers les speckles nucléaires et d’autres domaines nucléoplasmiques où ils participent à l’assemblage dynamique du spliceosome et à l’épissage des pré-ARNm.
Biogenèse et maturation de la télomérase
Les corps de Cajal participent à la maturation et au trafic intranucléaire du complexe télomérase, notamment par l’intermédiaire de WRAP53/TCAB1 (Telomerase Cajal Body Protein 1), qui se lie à la boîte CAB de l’ARN télomérase TERC et assure la localisation de la télomérase dans les corps de Cajal, ainsi que son adressage vers les télomères (Biogenesis and Regulation of Telomerase during Development and Cancer 2025).
La télomérase est une ribonucléoprotéine qui maintient la longueur des télomères en ajoutant des répétitions d’ADN télomérique aux extrémités des chromosomes, compensant la perte progressive de séquences lors des cycles de réplication (télomères et protection des extrémités chromosomiques).

La télomérase est étudiée dans le chapitre spécifique des télomères.
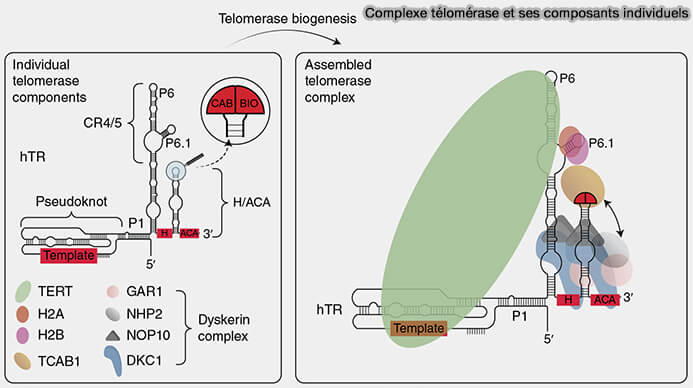
(Figure : vetopsy.fr d'après Chen et coll)
Retour à la classification des corps nucléaires
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauEnveloppe nucléairePores nucléairesLaminaCorps nucléairesTransport nucléocytoplasmiqueCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosqueletteFilaments d'actineFilaments intermédiairesMicrotubulesReproduction cellulaireBiochimieTransport membranaire Moteurs moléculairesVoies de signalisation