


Neurophysiologie : synapse
Vésicules synaptiques
Pools vésiculaires : modèle alternatif
- Neurophysiologie
- Cellules gliales et névroglie
- Neurones
- Potentiels membranaires
- Circuits neuronaux
- Neurotransmetteurs
- Récepteurs membranaires
- Synapses
{kind=link}

Le modèle des pools de vésicules synaptiques a ses limites et la relation entre vésicules synaptiques et système endolysosomal pourrait être d'un grand secours.
Le nombre ou la taille du pool de vésicules synaptiques est variable selon le type de neurones considéré, i.e. quelques dizaines pour les petites synapses telles que celles trouvées dans l'hippocampe.
Les pools de vésicules synaptiques (VS) peuvent être divisés en fonction :
- de leur déplacement par rapport à la membrane,
- leur relation temporelle entre la libération des vésicules et l'arrivée des potentiels d'action présynaptiques, cette dernière classification étant controversée.
Nouvelle approche des pools
La complexité toujours croissante de sous-populations de VS qui contribuent à des événements fonctionnels spécifiques de fusion ou de libération suggère une composition disparate du pool total de VS, avec des contributions potentiellement directes de différents organites du système endolysosomal ( composants communs des VS et du système endolysosomal).
composants communs des VS et du système endolysosomal).
1. Le modèle de pool des vésicules synaptiques (VS) a prouvé son utilité pour décrire la plasticité de la libération des neurotransmetteurs en termes de changements de tailles de pools VS (Nanoscale Remodeling of Functional Synaptic Vesicle Pools in Hebbian Plasticity 2020).
Le concept de pools de VS a été introduit en tant que modèle pour expliquer la diversité fonctionnelle des VS présentes dans le terminal présynaptique et leur contribution distincte aux différentes formes de libération des neurotransmetteurs (Synaptic Vesicle Pools and Dynamics 2012). .
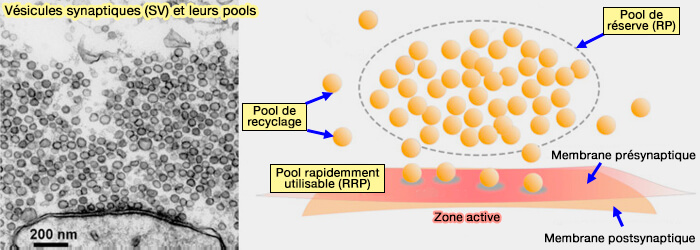
(Figure : vetopsy.fr d'après Zhang et coll)
2. Cependant, comme pour tout modèle, il a ses limites (Merits and Limitations of Vesicle Pool Models in View of Heterogeneous Populations of Synaptic Vesicles 2015).
Limites du modèle classique

Vous pouvez lire : Synaptic Vesicle Recycling and the Endolysosomal System: A Reappraisal of Form and Function (2022), article dont est tiré la plus grande partie de ce chapitre.
Pools
1. La principale limitation est que le modèle des pools reste trop déterministe dans son interprétation originale et, dans des cas spécifiques, il postule que des pools distincts avec une composition moléculaire spécifique contribuent à différentes formes de libération (Classification temporelle (modèle moléculaire)).
En réalité, le comportement dynamique des systèmes biologiques complexes, tels que le cycle des VS, est intrinsèquement stochastique.
- Les molécules sont distribuées au hasard à différentes populations de VS et des mécanismes de régulation complexes guident les résultats probabilistes.
- De ce fait, des résultats fonctionnels qualitativement similaires peuvent provenir des interactions d'un nombre différent et varié de molécules, chacune présente sur des VS en faible quantité absolue ( protéines des VS).
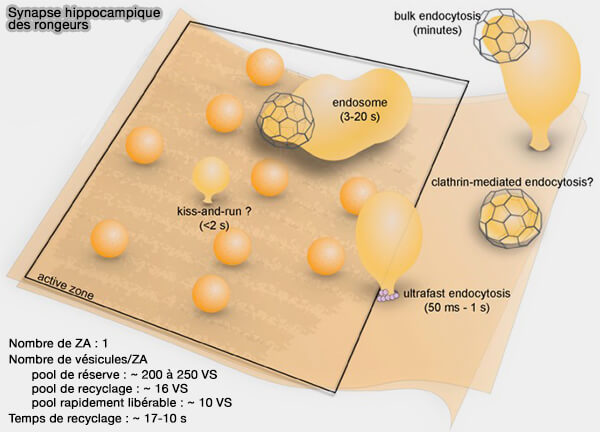
(Figure : vetopsy.fr d'après Gan et Watanabe)
2. Le modèle de pool n'explique pas non plus pourquoi le pool de repos in vitro et in vivo ne peut pas être libéré même après de longues périodes de stimulation synaptique (Synaptic Vesicle Pools and Dynamics 2012).
a. La réticences de ces VS à participer au recyclage dépendant de l'activité suggère qu'ils pourraient être des endosomes qui ont d'autres rôles que la contribution directe à la libération des neurotransmetteurs.
Par exemple, ces VS au repos pourraient servir de réservoir de molécules pouvant être incorporées dans le pool de recyclage à la demande, i.e. pendant le tri, pour permettre des changements plastiques de composition et de fonction des VS (The reserve pool of synaptic vesicles acts as a buffer for proteins involved in synaptic vesicle recycling 2011).
b. En outre, une partie du pool de repos peut en fait être composée d'une série d'intermédiaires endolysosomaux amorcés pour le transport vers les corps cellulaires neuronaux et les voies de dégradation.
(Figure : vetopsy.fr d'après Tagliatti et Glyvuc)
3. Dans de nombreux cas, le modèle de pool manque en fournissant une bonne corrélation entre la morphologie et la fonction.
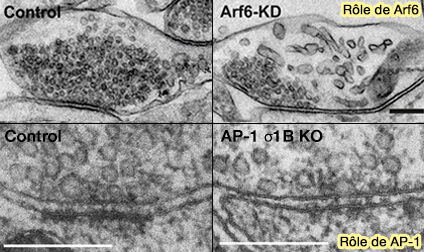
a. Ainsi, la perte de fonction des molécules, telles que la synaptobrévine 2/VAMP2, Munc13-1, Munc-18 et les canaux calciques, qui réduisent ou éliminent fortement les formes de libération des neurotransmetteurs, n'a pas d'impact majeur sur l'ultrastructure synaptique et le nombre de VS (Synapse and Active Zone Assembly in the Absence of Presynaptic Ca2+ Channels and Ca2+ Entry 2020).
b. En revanche, l'intégrité du regroupement des VS est compromise lorsque la fonction des protéines de tri endosomale clés est abrogée comme :
- AP-1 (AP-1/σ1B-adaptin mediates endosomal synaptic vesicle recycling, learning and memory 2010),
- Arf6 (Arf6 regulates the cycling and the readily releasable pool of synaptic vesicles at hippocampal synapse 2016).
4. Un support supplémentaire pour l'hypothèse que le pool de VS n'est pas un module autonome, fonctionnant indépendamment dans des terminaux synaptiques isolés, est fourni par l'existence d'un superpool de VS (Investigate Synaptic Vesicles Mobility in Neuronal Culture Axons by FRAP Imaging 2021).
Bien qu'il soit généralement admis que le pool de VS et le recyclage sont des phénomènes locaux, le pool ne se limite pas à une synapse particulière, mais couvre plusieurs synapses dans un seul axone (A Vesicle Superpool Spans Multiple Presynaptic Terminals in Hippocampal Neurons 2010).
Les VS et des protéines synaptiques en général se redistribuent rapidement entre les synapses voisines et même entre les synapses et les corps de cellules neuronales (Synaptic activity controls localization and function of CtBP1 via binding to Bassoon and Piccolo 2015).
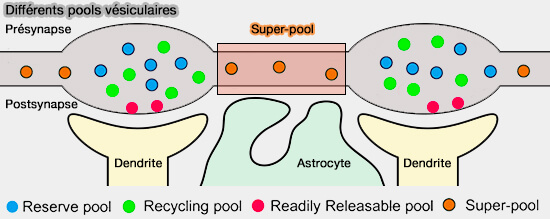
(Figure : vetopsy.fr d'après Zhang et coll)
5. Ainsi, le pool des VS est partagé entre de nombreuses synapses et se compose de plusieurs organites interconnectés qui se déplacent continuellement dans les deux sens entre les axones et les corps de cellules neuronales.

Les pools de VS et leur regroupement au niveau de la synapse sont pilotés par des programmes intrinsèques du neurone et le recyclage des VS n'est pas essentiel pour maintenir son intégrité.
Fusion vésiculaire
Contrairement au modèle des pools, la fusion des VS et la libération de neurotransmetteurs sont probablement une propriété émergente d'un petit sous-ensemble d'endolysosomes orchestrés dans la phase présynaptique qui a été sélectionnée par l'évolution pour soutenir la fonction centrale du système nerveux, la communication neuronale.
1. En effet, l'ultrastructure des terminaux présynaptiques excitateurs qui se sont développés en l'absence totale de libération de glutamate par l'expression de la toxine tétanique induite par Cre, a été largement préservée, avec un nombre normal de VS et a conduit à un affinement normal de la connectivité dans le cerveau en développement (Formation and Maintenance of Functional Spines in the Absence of Presynaptic Glutamate Release 2017).
2. Par ailleurs, les neurotransmetteurs excitateurs, i.e. glutamate et aspartate, et inhibiteurs, i.e. GABA et glycine sont des acides aminés.
- Le système endolysosomal est un site de stockage bien établi pour les acides aminés libres (Lysosomal metabolomics reveals V-ATPase and mTOR-dependent regulation of amino acid efflux from lysosomes 2017).
- Certains de ces acides aminés intraluménaux sont un élément clé d'un mécanisme endolysosomal central pour la détection des nutriments et le contrôle métabolique qui fonctionne dans pratiquement toutes les cellules et tous les organes.
3. En outre, le transport de neurotransmetteurs classiques et de neuromodulateurs vers les VS est piloté par des membres de la famille de transporteurs de soluté (SLC) qui sont largement exprimés dans diverses structures endolysosomales et sont essentiels à des processus vitaux au sein et en dehors du système nerveux.
- En plus de leur capacité à transporter des acides aminés, certains de ces canaux fonctionnent comme des canaux ioniques et régulent les gradients d'ions et l'acidification des endosomes (VGLUT1 functions as a glutamate/proton exchanger with chloride channel activity in hippocampal glutamatergic synapses 2017).
- L'importation de neurotransmetteurs pourrait donc être considérée comme un sous-produit de l'établissement de ces gradients électrochimiques, qui ont des rôles supplémentaires dans le fonctionnement des endosomes.
Conclusion

Les pools des VS peuvent être présentés comme une collection d'endosomes moléculairement hétérogènes et les VS supposées comme des intermédiaires de la voie endolysosomale.
1. En raison de l'accumulation d'ensembles spécifiques de molécules au cours du processus élaboré de tri endosomal, une petite fraction de ces endosomes s'engage dans des prises régulées (évoquées) ainsi que dans la libération constitutive (spontanée) dans les limites d'une synapse spécifique (Presynaptic origins of distinct modes of neurotransmitter release 2018).
2. Cependant, le cycle VS est imbriqué dans un cycle plus grand qui englobe les organites liés à la membrane qui résident dans les deux sites présynaptiques et dans des corps de cellules neuronales.
Cycle vésiculaire
NeurophysiologeCellules gliales et névroglieNeuronesPotentiels membranairesCircuits neuronauxNeurotransmetteursRécepteurs membranairesSynapsesStructure synaptiqueNeurotransmissionVésicules synaptiques (VS)Pools synaptiquesRôles du calcium dans l'électrosécrétionCycle synaptiqueTranslocation et attache (tethering)Amarrage (docking) et amorçage (priming)Fusion et exocytoseRecyclage des VSDifférents types d'endocytoseAcidification et remplissage des VSRegroupement des VSPlasticité et régulation synaptique