Dans le cycle des vésicules synaptiques, l'endocytose doit être couplée à l'exocytose pour pouvoir renouveler rapidement les vésicules fusionnées et maintenir la fonction synaptique et la stabilité structurelle de la zone active (ZA).
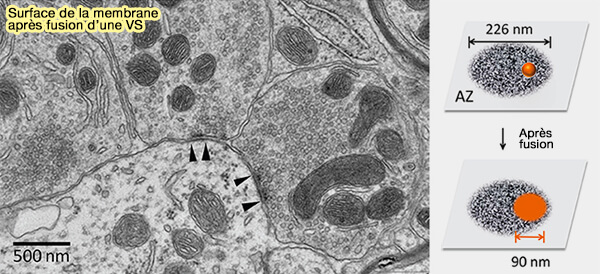
Compte tenu de la taille de la zone active (ZA), chaque fusion de VS peut considérablement augmenter la surface de la membrane plasmique de 6 362 nm2 et donc avoir un impact sur son ultrastructure et sa fonction.
Surface de la membrane après fusion d'une vésicule
(Figure : vetopsy.fr d'après Lou)
2. L'exocytose et l'endocytose des vésicules synaptiques (VS) sont étroitement liées dans le temps et dans l'espace lors du recyclage des VS, et ce processus est essentiel à la fonction synaptique et à la stabilité structurelle.
la taille du pool de VS, par la formation des stocks de vésicules,
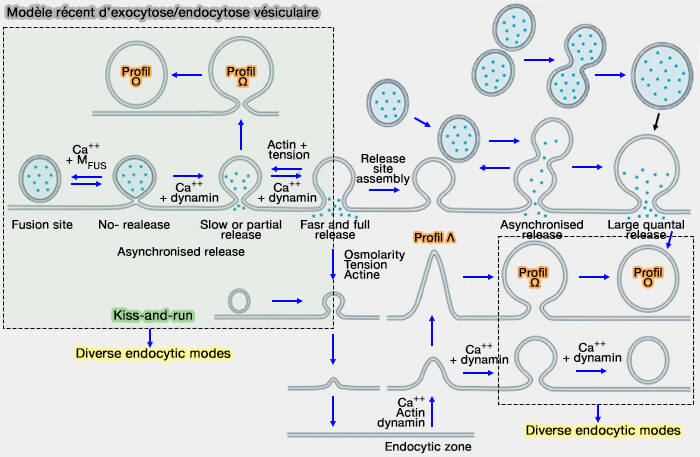
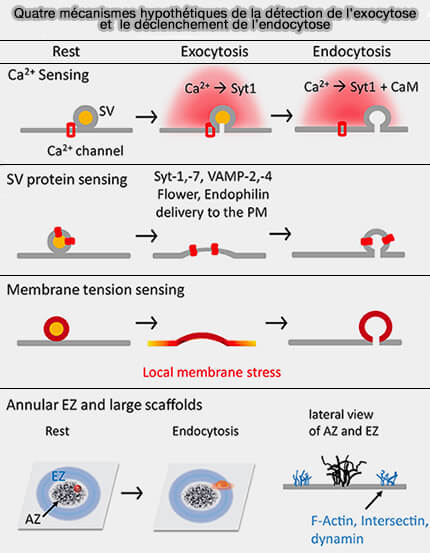
Quatre mécanismes possibles de couplage
exocytose/endocytose
(Figure : vetopsy.fr d'après Lou) le nettoyage, i.e. clairance, des sites de fusion encombrés par l’ensemble des protéines insérées dans la membrane vésiculaire pour rendre sa compétence au site de libération pour une nouvelle série de vésicules et pour éviter un ralentissement de l’exocytose (What is rate-limiting during sustained synaptic activity: vesicle supply or the availability of release sites 2010).
2. De plus, l’organisation spatiale des zones endocytaires adjacentes aux zones actives, i.e. les zones péri-actives, fournit une base structurelle pour un couplage efficace entre l’exocytose et l’endocytose du VS ( modèle par compression).
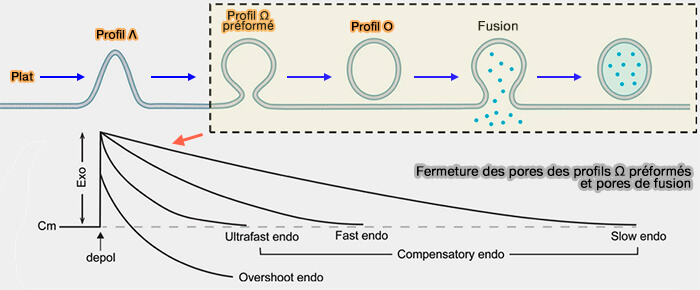
Transitions du profil de la vésicule d'endocytose
Vue d'ensemble
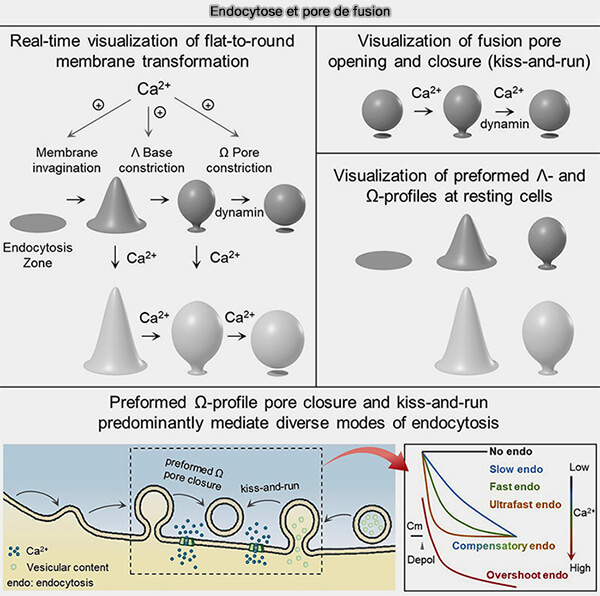
Dans ce chapitre, nous étudions les mécanismes sous-jacents à la transition Plat ➞ Λ ➞ Ω ➞ Ο observée en temps réel.
Les différences connues dans la machinerie moléculaire semblent difficiles à expliquer de manière satisfaisante les différents modes endocytaires, car la machinerie endocytaire est globalement identique.
1. Par contre, les études des transformations membranaires observées en temps réel offrent une explication solide pour la génération de modes endocytaires différents en termes de vitesses, de quantités et de tailles de vésicules avec la même machinerie moléculaire.
la transition préformée Λ ➞ Ω et la fermeture des pores de fusion (kiss-and-run),
la transition plat ➞ Λ ➞ Ω ➞ Ο.
b. L'endocytose excessive (endocytose > exocytose), overshoot endocytosis, est causée principalement par la transition préformée Ω ➞ Ο.
Fermeture des pores des profils Ω préformés et pores de fusion
(Figure : vetopsy.fr d'après Wu et Chan)
2. La différence notable entre ces différents modes est le courant calcique, déclencheur de chaque transformation de la membrane endocytaire et de chaque mode endocytaire.
Le courant calcique contrôle la vitesse de fermeture des pores des profils Ω préformés, le nombre et la taille des vésicules et ainsi, les afflux de calcium faibles à élevés génèrent
Endocytose et pores de fusion (Figure : vetopsy.fr d'après Shin et coll)
Le concept de profils Ω préformés pourrait expliquer plusieurs problématiques précédentes.
Comment une machinerie lente de transition plat ➞ Ο peut produire une endocytose rapide/ultrarapide ?
Pourquoi l'endocytose vésiculaire sécrétoire mesurée après la dépolarisation est beaucoup plus rapide que l'endocytose médiée par les récepteurs mesurée par l'ensemble de la transition Plat ➞ Ο ?
3. Par contre, la contribution de la transition plat ➞ O pourrait être plus significative si la transition plat ➞ Λ ➞ Ω ➞ O de faible probabilité est améliorée dans des conditions spécifiques ou dans des types de cellules.
Processus impliqués
Deux processus sont impliqués dans la plupart des modes endocytaires dits dynamine/actine-dépendants pour contribuer à la transition Plat ➞ Λ ➞ Ω ➞ Ο :



{kind=link}


 Ca++ et endocytoses),
Ca++ et endocytoses),