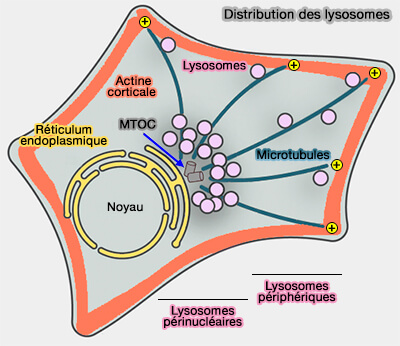
Les lysosomes, bien qu'ils puissent se déplacer de manière bidirectionnelle, sont localisés, à l'état d'équilibre, dans la région périnucléaire appelée MTOC (MicroTubule-Organizing Center) et sont appelés lysosomes périnucléaires.
1. Le réticulum endoplasmique (RE), le plus grand organite intracellulaire qui contient la majorité des lipides cellulaires, est étroitement associé à la biologie endosomale.
2. Les sites de contact RE/endosome participent à diverses fonctions moléculaires ( fonctions des MCS RE/endosomes), et en particulier au positionnement des lysosomes.
Par exemple, les MCS contrôlent le positionnement et la direction du transport liés :
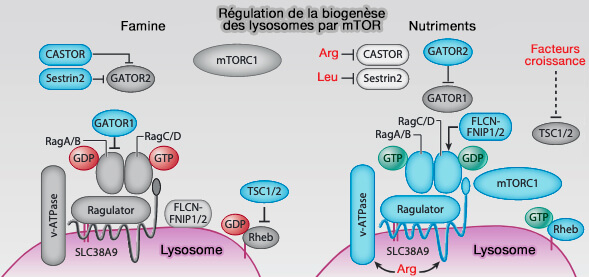
1. mTORC1 est un régulateur de croissance principal qui s'active du côté cytosolique des lysosomes en réponse aux nutriments, que ce soit lors de nutrition " normale " ou lors de famine ( autophagie) et ( voie mTORC1).
Régulation de la biogenèse des lysosomes par mTOR
(Figure : vetopsy.fr d'après Yang et Wang)
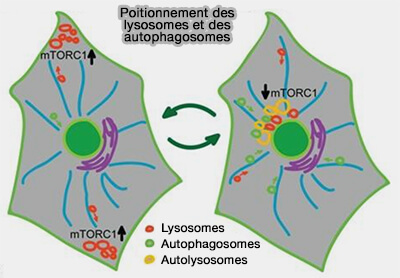
2. Le positionnement lysosomal est intimement associé à l'activité de mTORC1 et aux niveaux de nutriments, en particulier les niveaux d'acides aminés cytoplasmiques ou lysosomaux.
Positionnement des lysosomes et des autophagosomes
(Figure : vetopsy.fr d'après Korolchuk et coll)
la concentration de KIF2 et d'Arl8 diminue et les lysosomes périphériques se déplacent vers la région périnucléaire grâce à la dynéine,
la synthèse des autophagosomes augmente, et leur trafic et leur fusion avec les lysosomes s'effectuent dans la région périnucléaire ( fusion lysosomes/autophagosomes).
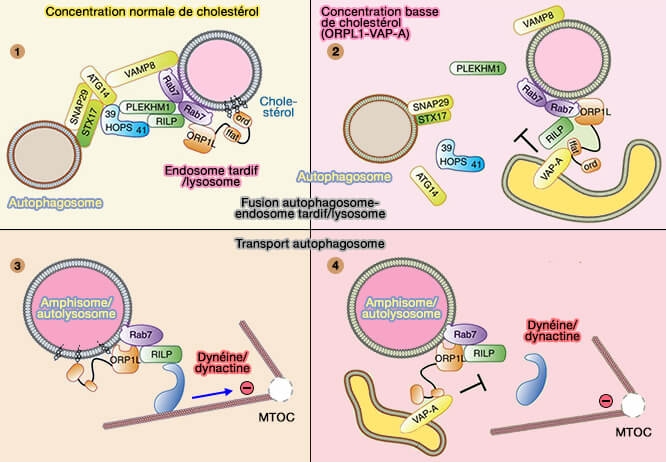
Modèle de ORPL1 pour la fusion et le trafic
(Figure : vetopsy.fr d'après Wijdeven et coll)
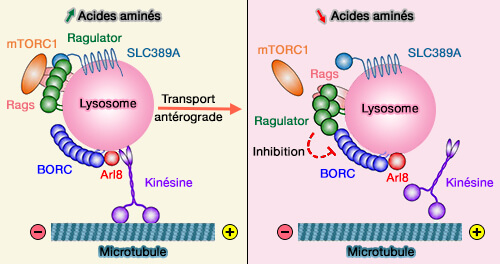
4. L'élimination de mTORC1 de la membrane lysosomale lors de famine suit l'inactivation du complexe Ragulator.
Ragulator est un complexe lysosomal qui contrôle l'activation de mTORC1 et l'autophagie en aval, en réponse à de faibles taux d'acides aminés.
BORC et Ragulator
(Figure : vetopsy.fr d'après Pu et coll)
Ragulator peut interagir avec le complexe BORC pour inactiver la diffusion endosomale tardive dépendante d'Arl8 par SKIP/kinésine 1.
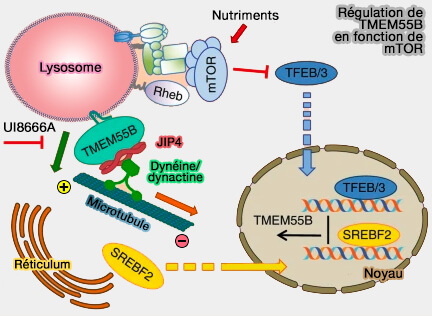
Régulation de TRM55B en fonction de mTOR
(Figure : vetopsy.fr d'après Willet et coll)
5. Après la famine, la protéine transmembranaire lysosomale TMEM55B, le PI-4 phosphatase transmembranaire, recrute l'adaptateur de dynéine JIP4, puis le complexe dynéine/dynactine sur les lysosomes, induisant le transport vers l'extrémité négative (-), i.e. vers MTOC.
La surexpression de TMEM55B provoque l'effondrement des lysosomes dans le centre cellulaire, tandis que l'épuisement de TMEM55B ou de JIP4 entraîne une dispersion vers la périphérie cellulaire.
La déplétion en TMEM55B ou JIP4 abolit le transport lysosomal rétrograde induit par la famine et empêche la fusion autophagosome-lysosome.
Une fois regroupés, les sites de contact membranaire formés entre les lysosomes et l'appareil de Golgi entraînent leur immobilisation dans la région périnucléaire par la folliculine - FLCN - ( MCS Golgi/lysosomes).
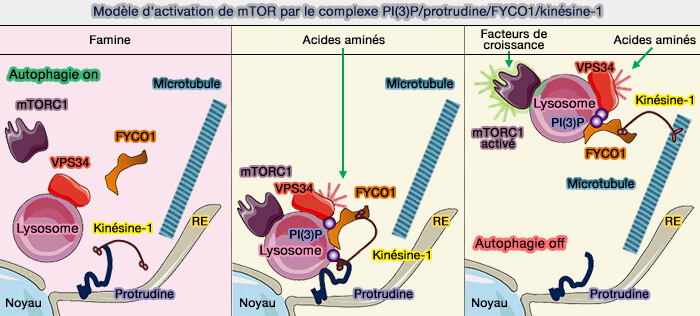
Les catégories d'endosomes sont caractérisées par des phosphoinositides différents qui vont aussi décider de leurs emplacements par leurs interactions avec les protéines motrices ou avec les différents adaptateurs ( endosomes et phosphoinositides).
Modèle d'activation de mTOR par le complexe PI(3)P/protrudine/FYCO1/kinésine-1
(Figure : vetopsy.fr d'après Hong et coll)
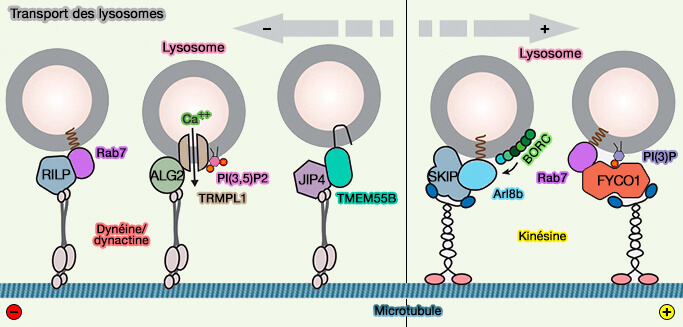
2. La maturation des endosomes tardifs en lysosomes s'accompagne d'une conversion de PI(3)P en PI(3,5)P2 généralement associée à la dégradation des cargos ( PI(3,5)P2 et endosomes).
Ce processus active le canal calcique lysosomal TRPML1 qui augmente les niveaux de calcium cytosolique pour recruter ALG2, le capteur de Ca++ cytosolique.
3. Le recrutement par les PIP nécessite le plus souvent une interaction de domaines protéiques solubles, PH, FYVE, PX… avec des groupes de tête lipidiques ( PIP et interactions endosomales) :
des Rabs ou des Arl/Arf et leurs effecteurs qui peuvent avoir des domaines interagissant avec les phosphoinositides qui modulent leurs interactions avec les protéines motrices,




 trafic des lysosomes) :
trafic des lysosomes) :