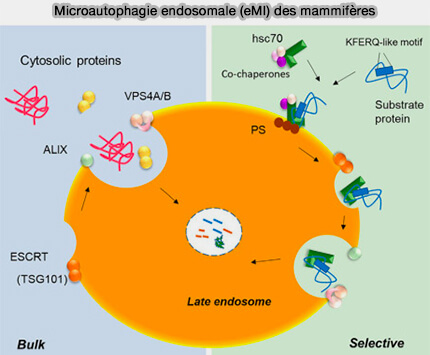
La microautophagie endosomale permet la séquestration sélective ou pas de protéines membranaires dans les endosomes sous forme de vésicules intraluminales (ILV) en vue de la fusion avec les lysosomes, alors que la microautophagie lysosomale peut directement dégrader certains organites et protéines transmembranaires.
b. Cependant, rapidement, une heure après l’induction de la dénutrition, ces récepteurs d’autophagie sont rapidement dégradés par un processus qui délivre sélectivement des récepteurs d’autophagie aux vésicules à l’intérieur des endosomes tardifs/corps multivésiculaires.
c. Ainsi, la privation en acides aminés provoque l’endocytose de récepteurs membranaires spécifiques, l’induction de la macroautophagie et la dégradation rapide des récepteurs de l’autophagie par la microautophagie endosomale.
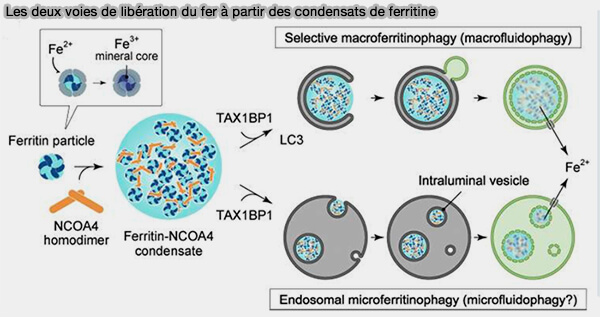
3. Au cours de la carence en fer, les particules de ferritine sont dégradées par les lysosomes pour la libération des ions Fe++ et ce par deux voies distinctes qui nécessitent NCOA4 ou Nuclear receptor coactivator 4 (The Role of NCOA4-Mediated Ferritinophagy in Ferroptosis 2021) :
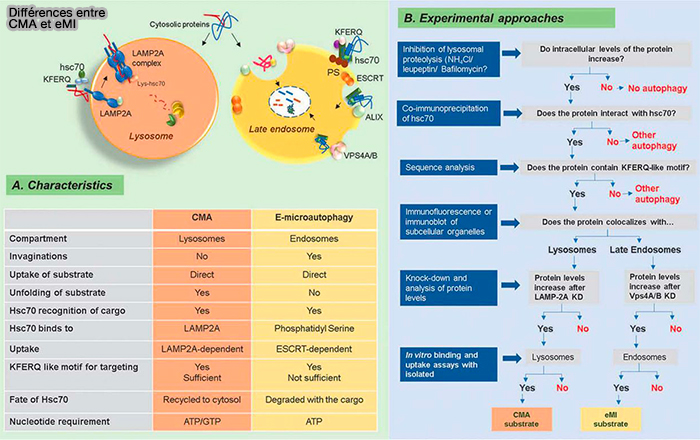
Chez D. melanogaster, qui manque d'autophagie à médiation par chaperon (CMA), la protéine HSC70 est recrutée via des interactions électrostatiques avec la phosphatidylsérine (PS), déformant la membrane endosomale, une nécessité pour l'autophagie.
La taille des lysosomes est généralement d'environ 500 nm de diamètre, ce qui est quatre à cinq fois plus petit que le diamètre des vacuoles dans la levure, ce qui rend difficile l'inspection de la membrane lysosomale invaginée.
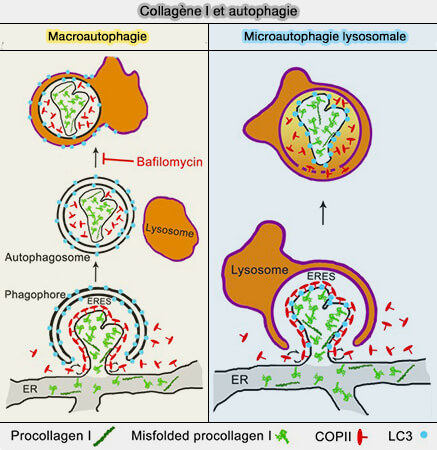
La microautophagie lysosomale peut dégrader certains organites et protéines transmembranaires dans la membrane lysosomale.
La microautophagie lysosomale peut être classée en fonction des substrats dégradés et est nommée, le cas échéant, comme la micro-x-phagie lysosomale, dans laquelle x indique les substrats dégradateurs.
On ne sait pas si le complexe ESCRT est impliqué dans ce processus.
Autres micro-x-phagies
D'autres organites peuvent subir cette microautophagie lysosomale.
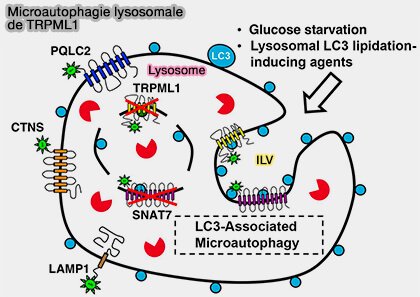
1. La membrane lysosomale et plusieurs protéines membranaires lysosomales, telles que TRPML1, peuvent être internalisées et dégradées dans la lumière lysosomale en cas de privation de glucose ou par un traitement qui induit un stress osmotique sur les lysosomes (Selective lysosome membrane turnover is induced by nutrient starvation 2020).
Microautophagie lysosomale de TRPML1
(Figure : vetopsy.fr d'après Lee et coll)
L’ionophoremonensine gonfle les lysosomes et, lors de la récupération, la taille des lysosomes est remodelée à la taille d’origine.
Ce processus dépend de la machine de base de la lipidation ATG, ATG3/ATG5/ATG7, mais pas ATG13, et on ne sait pas si le complexe ESCRT est impliqué dans ce processus.
Ce processus de remodelage est inhibé par la perte d’ATG5, mais pas d’ATG13, ce qui suggère un rôle de la microautophagie membranaire lysosomale dans le contrôle de la taille des lysosomes.
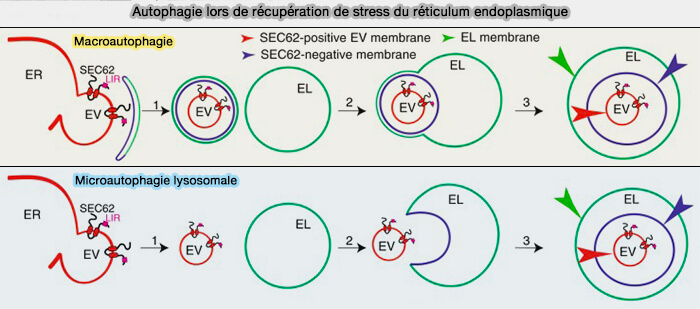
3. Dans la voie cGAS-STING, i.e. réponse immunitaire innée lors de la détection cytosolique d'un ADN à double brin, STING est dégradé par la microautophagie lysosomale ( STING et microautophagie lysosomale).
Ce processus est susceptible d'établir la structuration spatio-temporelle de la signalisation Wnt par la clairance lysosomale de Dickkopf1, l'antagoniste de Wnt.





 STING et microautophagie lysosomale).
STING et microautophagie lysosomale).