L'affinité de l'hémoglobine pour l'oxygène (O2) et le gaz carbonique (CO2) varie suivant plusieurs critères.
L’O2 et le CO2 sont des petites molécules non polaires qui traversent par diffusion simple, i.e. sans transporteur, ni énergie, la bicouche lipidique des membranes.
La coopérativité et la fonction physiologique de Hb sont fortement dépendantes de son affinité pour l'oxygène et de l'impact conséquent sur le pH de la solution.
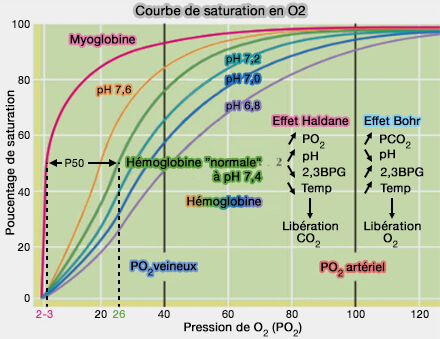
Courbes de saturation en O2 (myoglobine et hémoglobine)
(Figure : vetopsy.fr) Dans les poumons où la pression partielle en O2 (PO2) est élevée, le pH plus haut, la concentration en CO2 faible, et la tempérture plus basse par l'air inspiré, l’hémoglobine fixe facilement l’O2.
Dans les tissus où la PO2 est basse, la concentration en CO2 et en H+ est élevée et la température plus forte par le métabolisme, l’hémoglobine relâche l’O2.
L'effet Bohr désigne la diminution de l'affinité de l'hémoglobine pour l’O2 quand le pH diminue ou que la concentration en CO2 augmente.
Bien que cette réaction soit spontanée, elle se déroule très lentement en l’absence de catalyseur.
Or, le CO2 diffuse dans les hématies, et la réaction est considérablement accélérée par l’action de l'anhydrase carbonique de type II (CA-II), une enzyme très abondante dans ces cellules.
Cela provoque une diminution du pH du sang, ce qui favorise la dissociation de l'oxygène de l'hémoglobine, et permet aux tissus environnants d'obtenir suffisamment d'oxygène pour répondre à leurs besoins.
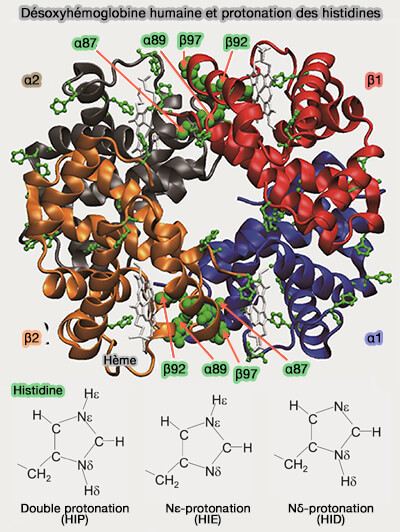
Désoxyhémoglobine humaine et protonation des histidines
(Figure : vetopsy.fr d'après Fujiwara et coll)
2. Lorsqu’elle libère l’O2 dans les tissus et forme la désoxyhémoglobine, la structure de l'Hb se modifie et devient plus basique.
a. La désoxyhémoglobine fixe les ions H+ sur les groupes imidazoles des histidines, i.e. 10 dans les sous-unités α et 9 dans les sous-unités β2.
À l'état T, la valeur du pKa, i.e. pH auquel un groupement acide est à moitié dissocié, de leur groupement imidazole augmente, i.e. pKa ≈ 7.8.
b. Les résidus peuvent adopter trois états de protonation, i.e. double protonation (HIP), Nε-protonation (HIE) et Nδ-protonation (HID).
c. Ce processus stabilise la forme T de l’hémoglobine, la forme à faible affinité pour l’O2, i.e. l'O2 est facilement libéré dans les tissus.
En fixant les ions H+ libérés par la réaction du CO2, l’hémoglobine maintient le pH sanguin autour de 7,4 malgré la production constante de CO2 et d’acides dans l’organisme.
Remarque : contrairement à HbO2 qui est diamagnétique, i.e. ne contient pas d'électron non apparié, i.e. Fe++ lié à 2 atomes d'oxygène, la désoxyhémoglobine comporte 4 atomes de Fe++ avec chacun quatre électrons non appariés, ce qui lui confère des propriétés paramagnétiques utilisées dans l'imagerie par résonance magnétique
fonctionnelle (IRMf).
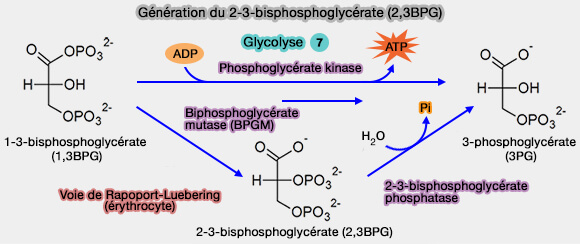
3. L'effet Bohr a aussi comme facteur le 2,3-bisphosphoglycérate (2,3-BPG) produit dans les érythrocytes matures dépourvus de mitochondries par une voie dérivée de la glycolyse, appelée shunt du 2,3-BPG ou voie de Rapoport-Luebering.
a. Dans la glycolyse classique, le 1,3-bisphosphoglycérate (1,3BPG), produit par l'étape 6, forme à l'étape 7 du 3-phospho-D-glycérate (3PG) par la phosphoglycérate kinase qui récupère des ATP.
Le 3-phospho-D-glycérate (3PG) sera isomérisé par l'étape 8 en 2-phospho-D-glycérate (2PG) par la phosphoglycérate mutase (PGM), i.e. EC 5.4.2.11, qui nécessite par ailleurs l'intermédiaire du 2,3-BPG.
b. Dans la voie de Rapoport-Luebering, la bisphosphoglycérate mutase (BPGM)(EC 5.4.2.4), spécifique des érythrocytes et des cellules placentaires, transforme le 1,3BPG en 2,3BPG, en shuntant l'étape 7.
Le 2,3BPG peut ensuite être converti en 3PG par la bisphosphoglycérate phosphatase (EC 3.1.3.13) pour réintégrer la glycolyse.
Génération du 2-3-biphosphoglycérate (2,3BPG)
(Figure : vetopsy.fr)
c. Le 2,3BPG peut se loger dans la cavité centrale de la désoxyhémoglobine, i.e. forme T, qui est plus large que la forme R, en se liant aux sous-unités β.
Le 2,3-BPG est un composé fortement chargé négativement par ses deux groupements phosphate.
Il interagit avec des résidus positifs principalement, Histidine β143, Lysine β82, Histidine β2 et Valine β1 N-terminale par des liaisons ioniques et des ponts hydrogène.
Remarque : en altitude, le taux de 2,3-BPG augmente et permet une meilleure libération d’oxygène dans les tissus, malgré une PO2 réduite.
Les résidus de liaison de Band 3 chevauchent partiellement ceux de 2,3-BPG, mais certains diffèrent, i.e. Lysβ82, Hisβ2, Hisβ143, Lyβ127, mais aussi Valβ1 (N-terminal)
a. Elle fait partie des échangeurs anioniques qui régulent l'échange HCO3-/Cl- (bicarbonate/chlore) à travers la membrane érythrocytaire (Band 3 et érythrocytes).
Dans les tissus, l'augmentation de la concentration de bicarbonate dans l'érythrocyte provoque son transport par AE1 dans le plasma sanguin par un échange électroniquement neutre avec les ions chlorure (Cl-).
Cette réaction provoque l'accumulation de protons et la baisse du pH, pour la libération de l'oxygène.
b. En outre, elle déplace les enzymes glycolytiques comme la GAPDH, augmentant la production de 2,3BPG qui favorise la libération d’O2.
En présence d'oxyhémoglobine, Band 3 est lié, par son extrémité N-terminale, aux enzymes glycolytiques qui sont de ce fait moins actifs.
En présence de désoxyhémoglobine, elle s'y attache par sa région N-terminale et libère ainsi les enzymes glycolytiques dans le cytosol pour augmenter la production de 2,3BPG.
Remarque : des espèces d'oiseaux, de reptiles et de poissons utilisent l'ATP à la place du 2,3BPG.
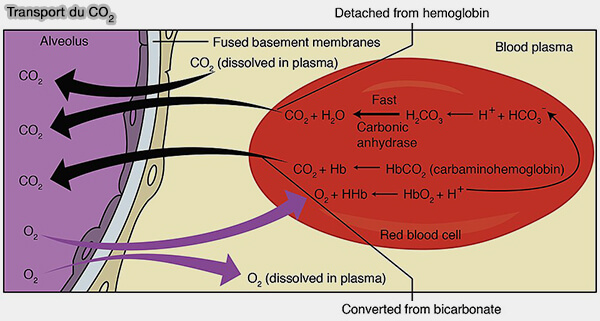
CO2 peut être transporté de 3 façons vers les poumons :
sous forme de bicarbonate HCO₃⁻ (~70 %) dans le sang,
sous forme de carbaminohémoglobine (~20–25 %) dans les érythrocytes,
dissous dans le plasma (~7 %).
1. Le CO2 est transporté majoritairement sous forme de bicarbonate $\ce{HCO3-}$ vers les poumons dans lesquels la pression partielle en O2 (PO2) est élevée, le pH plus haut et la concentration en CO2 faible.
2. Le CO2 peut se fixer sur l'hémoglobine, mais, contrairement à l'O2, il ne se fixe par sur l'hème, mais uniquement sur des groupes amines libres (-NH2) qui semblent dépendre du contexte :
des groupes α-aminés, au niveau du squelette principal de la protéine, comme les valines dans l'HbA, situés à l’extrémité N-terminale de chaque chaîne protéique à la surface de la protéine,
des groupes ε-aminés, i.e. situés sur la chaîne latérale, de certains résidus de lysine (et parfois d'autres acides aminés basiques).
La fixation du CO2 forme une carbaminohémoglobine, i.e. NH-COO- est un carbamate.
Cette réaction libère des ions H+ qui acidifient le sang qui, comme précédemment, stabilise la forme T de l’hémoglobine et favorise la libération de l’O2.
John Scott Haldane (1860-1936)
En même temps, par l'effet Haldane, inverse de l'effet Bohr, la désoxyhémoglobine se lie plus facilement au CO2 que l’oxyhémoglobine, ce qui permet son transport vers les poumons.
John Scott Haldane (1860-1936) est un physiologiste écossais qui découvrit cet effet en expérimentant, de manière surprenante, des cooktails de gaz potentiellement mortels sur son fils et lui-même, tout en enregistrant leur effet sur son esprit et son corps (Diving For Science 2009, p: 112).
Remarque : les effets Haldane favorisent :
l’absorption du CO2 par l’Hb dans les tissus périphériques où elle libère de l’oxygène vers le tissu,
la libération de CO2 de l’Hb dans les poumons où l’oxygène de l’air inspiré se lie à nouveau à l’Hb.
3. Le CO2 dissous circule simplement dans le plasma, sans transformation chimique, car il est 20 fois plus soluble dans l'eau que l'O2.
Dans les poumons, le CO2 dissous diffuse directement des capillaires vers les alvéoles pour être exhalé.
Le CO2 dissous est mesuré dans les gaz du sang artériel (PaCO2), d'environ ≈ 40 mmHg.
Si la PCO2 est augmentée, l'hypercapnie provoque une acidose respiratoire.
Si la PCO2 est basse, l'hypocapnie provoque une alcalose respiratoire.
Dans les poumons : recharge en O2 et élimination du CO2
Dans les poumons où la ression partielle en O2 (PO2) est élevée, le pH plus haut, la concentration en CO2 faible et la température plus basse par l'air inspiré, l’hémoglobine fixe facilement l’O2.
1. L’hémoglobine reprend sa forme R, la forme à haute affinité pour l’O2, se recharge en O2 et libère les ions H+.
$\ce{HHb}$ $\longrightarrow$ $\ce{Hb- + H+}$
2. Ces ions H+ interagissent avec les deux premières voies de transport du CO2 pour son élimination (effet Haldane).
a. Ils sont recombinés avec les bicarbonates $\ce{HCO3-}$ pour régénérer du CO2.
3. Avec le CO2 dissous, i.e. la troisième voie de transport, le CO2 diffuse directement des capillaires vers les alvéoles pour être exhalé car la pression partielle du CO2 (PCO2) du sang est supérieure à la PCO2 des alvéoles pulmonaires.
Ce processus passif et rapide est favorisé par la surface, 70 à 100 m2, et la finesse des alvéoles, 0,2 µm, entourées d’un réseau dense de capillaires sanguins.





 interaction de Band 3 avec l'hémoglobine).
interaction de Band 3 avec l'hémoglobine).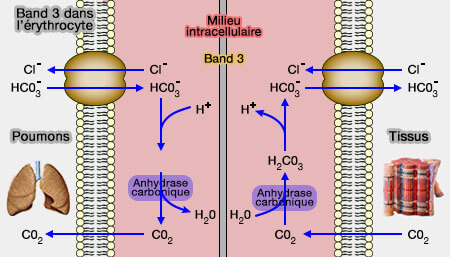
{kind=link}
