


Neurophysiologie : synapse
Vésicules synaptiques : cycle des vésicules synaptiques
3-4. Amarrage (docking) et amorçage (priming)
Processus mis en jeu et différents modèles
- Neurophysiologie
- Cellules gliales et névroglie
- Neurones
- Potentiels membranaires
- Circuits neuronaux
- Neurotransmetteurs
- Récepteurs membranaires
- Synapses
{kind=link}

Différents modèles essaient d'expliquer l'amarrage complexe des vésicules et les recherches sont encore en cours pour une meilleure compréhension de la composition du pool rapidement libérable (RRP) et de ses différentes composantes.
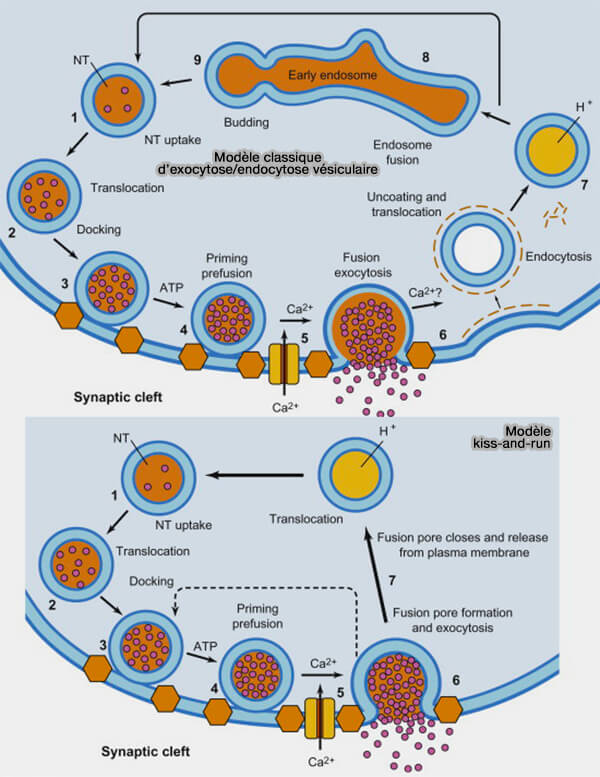
(Figure : vetopsy.fr d'après Pigino et coll)
La libération de vésicules synaptiques (VS) est régulée par la concentration de calcium dans la terminaison présynaptique ( afflux de Ca++).
afflux de Ca++).
- Avant sa libération, une vésicule doit se lier à une structure spécifique au sein de la zone active (ZA), le site d'amarrage (DS ou Docking Site).
- L'amarrage (docking) de la vésicule est dépendant du Ca++.
Un modèle simple pourrait ne pas suffire dans toutes les conditions, car, en pratique, plusieurs protéines pourraient participer aux transitions dépendantes du calcium.

Vous pouvez lire la thèse de Melissa Silva Medina Weil qui inspire fortement ce chapitre: The last steps before synaptic vesicle release : calcium-dependent docking and functional implication of docking site organisation (2024).
Modèle sans pool et sans site d'amarrage
Dans ce modèle, les vésicules synaptiques (VS) seraient libérées à partir de n'importe quel point de la membrane, sans pools distincts de VS.
1. L'hétérogénéité du temps de libération a été modélisée comme la distance entre les Cav/VGCC et les VS.
Cette distance est directement liée à la probabilité de libération et est utilisée pour expliquer la composante rapide et lente, ainsi que les modèles de plasticité (Loose Coupling Between Ca2+ Channels and Release Sensors at a Plastic Hippocampal Synapse 2014).
2. Bien que certains modèles ne précisent pas comment les VS restent à un endroit déterminé avant d'être relâchés, d'autres incluent des attaches entre Cav/VGCC et VS (Nanoscale Distribution of Presynaptic Ca2+ Channels and Its Impact on Vesicular Release during Development 2015).
Modèles avec pools et sites d'amarrage
L'approche des VS à la membrane peut se réaliser de deux façons différentes.
1. Dans les modèles séquentiels, les VS suivent une série d'étapes avant d'être prêtes à être libérées et, une fois amarrées, les VS sont libérées avec une probabilité pr ( occupation du site d'amarrage).
Des modèles séquentiels ont été utilisés pour expliquer la variance et la distribution des libérations ainsi que la plasticité entre différentes préparations. Les articles suivants apportent des précisions sur ce modèle.
- Fully-primed slowly-recovering vesicles mediate presynaptic LTP at neocortical neurons (2023)
- Increased vesicle fusion competence underlies long-term potentiation at hippocampal mossy fiber synapses (2023)
- A sequential two-step priming scheme reproduces diversity in synaptic strength and short-term plasticity (2022)
- Quantal analysis estimates docking site occupancy determining short-term depression at hippocampal glutamatergic synapses (2021)
- Rapid regulation of vesicle priming explains synaptic facilitation despite heterogeneous vesicle:Ca2+ channel distances (2020)
- Direct imaging of rapid tethering of synaptic vesicles accompanying exocytosis at a fast central synapse (2020).
2. Dans les modèles parallèles, la libération se produit à partir de deux populations différentes de VS, qui sont libérées avec des probabilités différentes, pr1 et pr2 (Bassoon Speeds Vesicle Reloading at a Central Excitatory Synapse 2010).
Ces modèles parallèles pourraient expliquer certains faits dans des préparations particulières comme par exemple :
- les terminaisons de l'habenula médiane (MhB) sur le noyau interpédonculaire ou IPN ( pool supplémentaire ?),
- les terminaisons des fibres moussues du cervelet sur les cellules granulaires post-synaptiques (GC) de la souris (Calcium dependence of neurotransmitter release at a high fidelity synapse 2021).
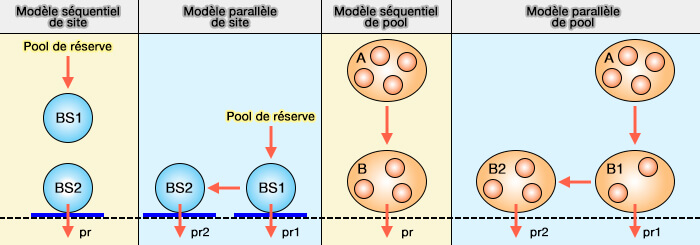
(Figure : vetopsy.fr d'après Weil)
3. La figure ci-dessus représente les modèles de site et de pools vésiculaires.
a. Un modèle de site séquentiel consiste en des sites d'amarrage fonctionnant en parallèle dans une zone de disponibilité.
- BS1 et BS2, i.e. BS ou Binding Site, représentent respectivement le site de liaison distal et le site de liaison et un site proximal qui peuvent être vides ou pleins.
- Les VS du pool de réserve transitent vers BS1 puis vers BS2, d'où ils sont libérés avec une probabilité pr.
b. La différence avec un modèle de site parallèle (B) est que :
- les VS peuvent également être libérés de BS1,
- BS1 et BS2 ont une probabilité de libération différente, i.e. pr1 et pr2.
c. Les modèles de pools sont modélisés en fonction de leur volume et de leur état de remplissage.
- Les pools séquentiels et parallèles sont modélisés de la même manière que précédemment.
- Dans les pools parallèles, le pool rapidement libérable (RRP) correspond à la somme de B1 et B2.
3. Lors de la comparaison directe des modèles de pool parallèle et séquentiel dans les cellules chromaffines, le modèle séquentiel décrirait mieux les expériences et peut être facilement ajusté avec les connaissances actuelles pour tenir compte des mutations (A Sequential Vesicle Pool Model with a Single Release Sensor and a Ca2+-Dependent Priming Catalyst Effectively Explains Ca2+-Dependent Properties of Neurosecretion 2013).
- Cette comparaison de modèles a incorporé deux ensembles différents de VS qui étaient auparavant envisagés comme des pools.
- Toutefois, dans ces modèles, les sites de libération n'entrent pas en ligne de compte.
Alternative aux modèles précédents
Les deux composantes de libération évoquées, i.e. synchrone et asynchrone, ont fourni de bonnes preuves de l'existence d'un ensemble hétérogène de vésicules synaptiques (VS).

Ces deux composantes peuvent être modélisées comme différentes étapes ou états d'amarrage.
1. Deux modèles de sites séquentiels peuvent être envisagés et supposent que les sites d'amarrage fonctionnent en parallèle avec deux étapes séquentielles de liaison par site d'amarrage.
- le modèle RS/DS ((Remplacement Site/Docking Site),
- le modèle LS/TS (Loose State/Tight State).
La transition entre les deux étapes de liaison est rapide et la libération ne se fait qu'à partir du site le plus proche de la membrane (DS ou LS).
1. Même au repos, les états amarrés et amorcés des VS peuvent être labiles et très dynamiques et peuvent fluctuer entre (Munc13-1 and Munc18-1 together prevent NSF-dependent de-priming of synaptic vesicles 2017 et Trans-SNARE complex dynamics and number determine nascent fusion pore properties 2018) :
- un état lâchement amarré et amorcé (LS ou loosely docked and primed state) dans lequel les complexes SNARE ne sont que partiellement zippés,
- un état étroitement amarré et amorcé (TS ou tightly docked and primed), dans lequel la fermeture éclair s'est refermée sur une plus grande distance.
En effet, les états distincts d'amarrage membranaire des VS sont déterminés par les complexes SNARE et un ensemble défini de protéines d'amorçage des VS régulant les SNARE ( protéines impliquées).
Les complexes SNARE peuvent exister dans un équilibre dynamique entre un état lâche et serré, et la conversion entre les états LS et TS des VS semble être extrêmement rapide lors d'une activité synaptique à haute fréquence soutenue et d'augmentations concomitantes de Ca++.
2. Dans les circonstances testées, il existe un échange dynamique dépendant du Ca++ entre les deux états des VS étroitement et lâchement ancrés à la membrane de la zone active (ZA).
- Dans les synapses sauvages (WT), l'équilibre entre les états LS et TS n'est presque jamais complètement déplacé vers TS.
- Ainsi, une stabilisation transitoire du TS, ou même un retour au LS, peut également se produire dans des conditions physiologiques normales.
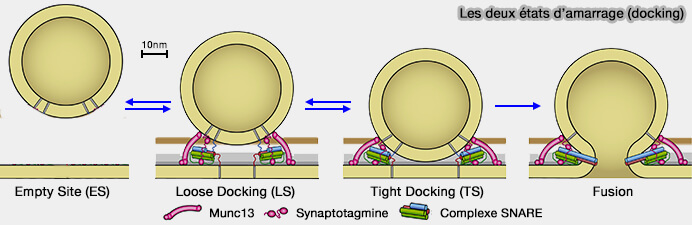
(Figure : vetopsy.fr d'après Neher et Brose)
Dans la figure ci-dessus, les VS, les membranes et les protéines sont représentées à peu près à la même échelle.
La VS, i.e. 45 nm de diamètre, est placée à 8 nm de distance membrane à membrane, distance à laquelle les protéines SNARE commencent à interagir par des forces (Energetics and dynamics of SNAREpin folding across lipid bilayers 2007) ou forment un complexe lâche dans les études sur les liposomes (Arrest of trans-SNARE zippering uncovers loosely and tightly docked intermediates in membrane fusion 2018).
- L'aire en gris foncé correspond à la zone d'amarrage/amorçage serrée telle que définie par Imig (The Morphological and Molecular Nature of Synaptic Vesicle Priming at Presynaptic Active Zones 2014).
L'aire en brun est la zone de densité VS accrue chez les mutants défectueux pour l'amarrage/amorçage.
- L'aire en gris clair correspond à la zone d'amarrage/amorçage serrée telle que définie par Chang (Synaptotagmin-1 drives synchronous Ca2+ triggered fusion by C2B domain-mediated synaptic vesicle-membrane attachment 2018).
Modèles RS/DS et LS/TS
NeurophysiologeCellules gliales et névroglieNeuronesPotentiels membranairesCircuits neuronauxNeurotransmetteursRécepteurs membranairesSynapsesStructure synaptiqueNeurotransmissionVésicules synaptiques (VS)Pools synaptiquesRôles du calcium dans l'électrosécrétionCycle synaptiqueTranslocation et attache (tethering)Amarrage (docking) et amorçage (priming)Fusion et exocytoseRecyclage des VSDifférents types d'endocytoseAcidification et remplissage des VSRegroupement des VSPlasticité et régulation synaptique