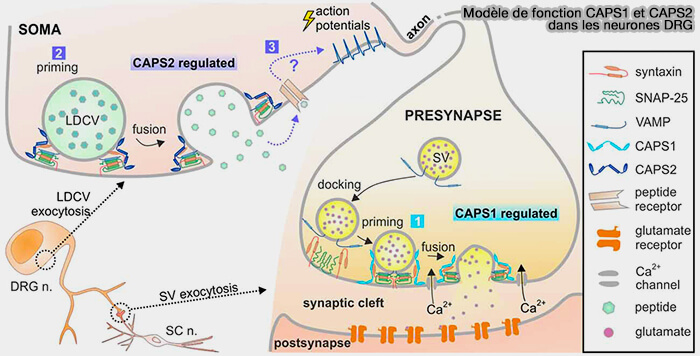
La libération de vésicules synaptiques (VS) est régulée par la concentration de calcium dans la terminaison présynaptique qui est évaluée par les senseurs calciques.
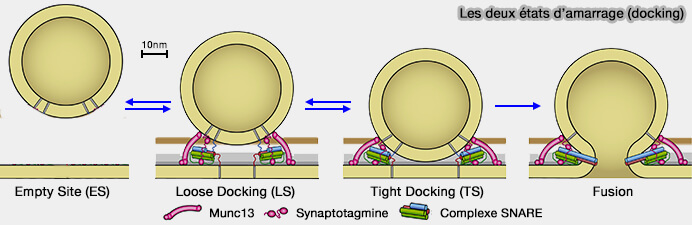
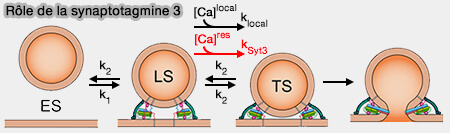
Les deux états d'amarrage LS et DS
(Figure : vetopsy.fr d'après Neher et Brose)
Dans le cycle vésiculaire, les senseurs calciques sont impliqués dans :
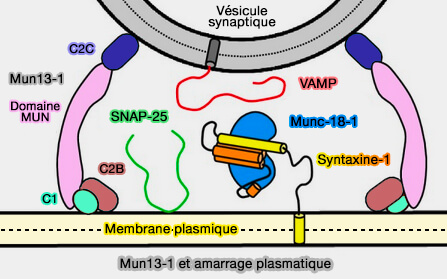
a. Munc13-1 est essentiel à l'amarrage (docking), à l'amorçage (priming), à la fusion des vésicules synaptiques (VS) et à la libération des neurotransmetteurs par son implication comme chaperon du complexe SNARE avec les protéines Sec1/Munc18 (SM).
Munc-13 augmenterait l'amarrage proximal, ce qui est cohérent avec les théories antérieures sur son rôle dans l'équilibre des pools vésiculaires au sein du pool rapidement libérable (RRP) qui favorise un état de probabilité de libération élevée.
Munc13 ne contrôle pas seulement l'amorçage de base du VS dans l'état LS, mais peut également être utilisée par les neurones de manière ciblée pour contrôler les transitions LS ➞ TS dépendantes de l'activité.
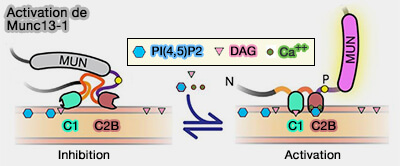
3. La signalisation Ca++/calmoduline, le Ca++/PI(4,5)P2 et le diaycylglycérol (DAG) peuvent conduire à des changements conformationnels dans la structure des Munc13 liés aux complexes SNARE qui favorisent la progression vers l’état TS.
Les interactions allostériques au sein de l'ensemble de l'appareil de fusion VS peuvent combiner les effets régulateurs des Munc13 avec ceux des synaptotagmines pour générer un STP dépendant de l'activité sur une large échelle de temps et une large plage de [Ca2+]i.
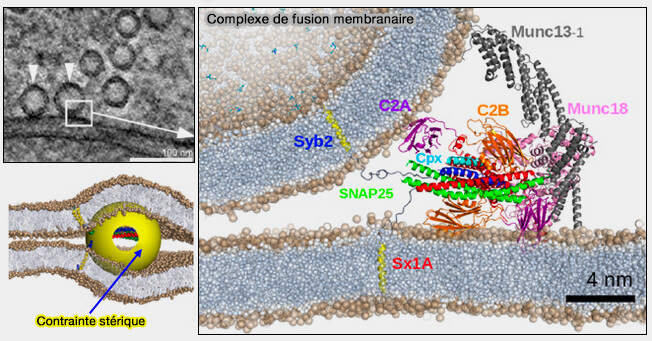
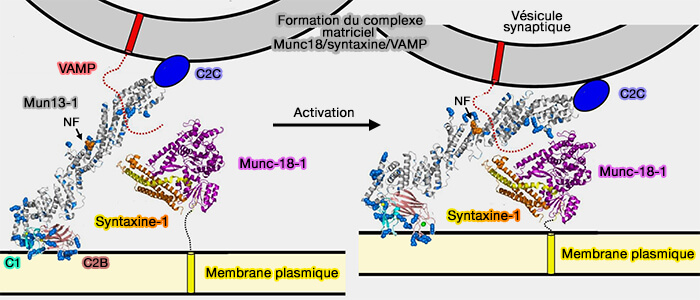
Formation du complexe matriciel Munc18/syntaxine/VAMP
(Figure : vetopsy.fr d'après Xu et coll)
a. Les synapses chez les souris KO de Syt3 présentent une dépression synaptique à court terme accrue, et la récupération est plus lente et insensible au Ca++ présynaptique résiduel.
Cet effet peut être modélisé comme une augmentation transitoire dépendante du calcium de l'amarrage proximal.
Syt3 serait en partie responsable du taux plus rapide observé aux hautes fréquences, mais un deuxième amplificateur avec les caractéristiques de Munc13 est nécessaire pour s'adapter à toutes les données.
b. Syt3 sert également à la facilitation synaptique, dans des conditions de faible probabilité de libération des vésicules.
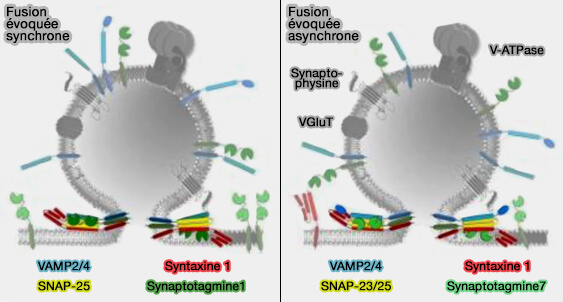
b. Même dans les neurones contenant la synaptotagmine-1, l'ablation de la synaptotagmine-7 a partiellement altéré la libération asynchrone induite par des trains prolongés de stimulus à haute fréquence.
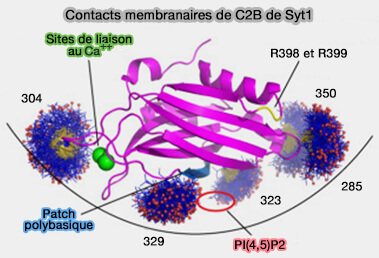
La fonction de la synaptotagmine-7 nécessite sélectivement ses sites de liaison au Ca++ du domaine C2A, alors que la fonction de la synaptotagmine-1 nécessite ses sites de liaison au Ca++ du domaine C2B.
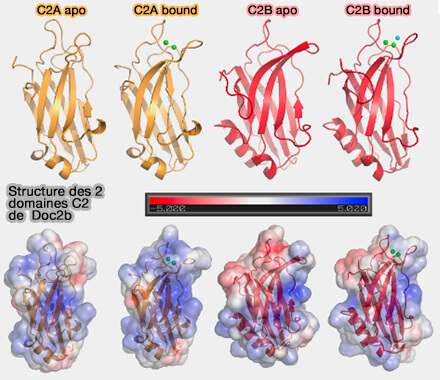
La régulation à la hausse et à la baisse des niveaux d'expression de Doc2 dans les neurones de l'hippocampe a augmenté ou diminué, respectivement, la phase lente de la transmission synaptique.
b. Lors de KO de Doc2, on observe une augmentation du nombre de VS amarrées à 11 ms, de la durée de l'amarrage transitoire et un état d'équilibre plus élevé que celui des souches sauvages.
Un double KO de Doc2 et de Syt7 obtient le même résultat.
Syt-7 pourrait agir comme capteur pour l'amarrage transitoire qui alimente les VS à Syt1 pour une libération synchrone ou à Doc2 pour libération asynchrone, mais l'interprétation du point de vue du modèle RS/DS.
Dans le modèle LS/TS, Syt7 pourrait être responsable de la transition LS ➞ TS tandis que Syt1 et Doc2 participeraient à l'étape de fusion.
1. Toutefois, lors de déficience de Syt1 synaptique, la libération des neurotransmetteurs est désynchronisée.
Interactions synaptostagmine/SNARE
(Figure : vetopsy.fr d'après Nyenhuis et coll)
a. Cette désynchronisation peut être partiellement restaurée par des mutants Syt1 qui n'ont pas la capacité de se lier aux membranes et/ou au complexe SNARE.
Contrairement au sauvetage avec le Syt1 de type sauvage (WT), ces variantes mutantes de Syt1 ne supportent la libération synchrone des VS que pour les potentiels d'action (PA) qui sont précédés par un autre PA dans une fenêtre temporelle de 10 à 50 ms.
Le nombre de VS à une distance de -5 nm de la membrane plasmique est augmenté à moins de 10 ms après un premier PA, mais rediminue ensuite.
Ainsi, ce n'est que dans une fenêtre temporelle de 10 à 50 ms que la distribution spatiale des VS ressemble à celle des synapses exprimant Syt1 WT.
déclenchement du processus de fusion en réponse à un PA.
Un échange dynamique dépendant du Ca++ entre deux états VS amorcés, i.e. étroitement (TS) et lâchement (LS) ancrés à la membrane de la ZA ( vésicules LS et TS).
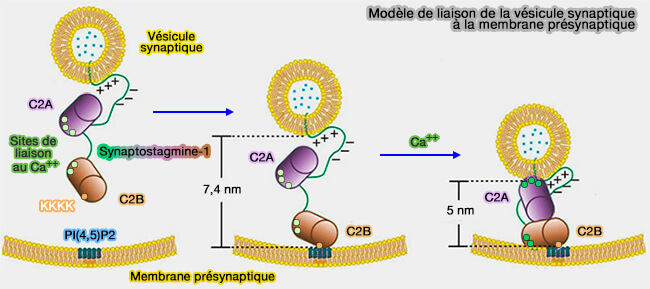
Modèle de liaison de la vésicule synaptique à la membrane présynaptique
(Figure : vetopsy.fr d'après Lin et coll)



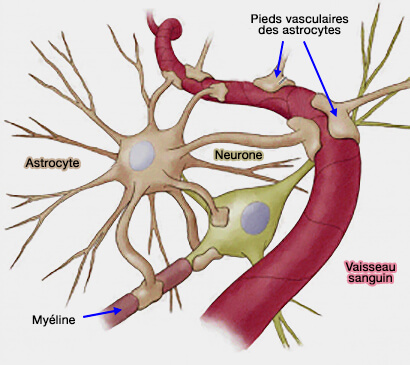{kind=link}



 Doc2 et Syt1 dans la libération spontanée) et l'endocytose ultrarapide (UFE).
Doc2 et Syt1 dans la libération spontanée) et l'endocytose ultrarapide (UFE).