


Trafic vésiculaire
Endocytose clathrine-indépendante (CIE) à grande échelle
Phagocytose : récepteurs phagocytaires
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Transport membranaire
- Transports membranaires sans mouvements membranaires
- Transports membranaires avec mouvements membranaires : trafic vésiculaire
- Protéines membranaires
- Moteurs moléculaires
- Voies de signalisation

Les phagocytes sont confrontés à un large éventail de cibles et de ligands phagocytaires, ce qui nécessite l'expression d'un vaste répertoire de récepteurs membranaires divisés en récepteurs opsoniques ou non opsoniques.
Récepteurs phagocytaires

Les récepteurs et leur détection sont détaillés dans : Phagocytosis: Our Current Understanding of a Universal Biological Process (2020) et Diffusion Barriers, Mechanical Forces, and the Biophysics of Phagocytosis (2016).
Récepteurs non opsoniques
Les récepteurs non opsoniques identifient directement des motifs moléculaires distincts sur la particule à ingérer.
1. Les lectines sont des protéines qui se lient spécifiquement et de façon réversible à certains glucides, en particulier les lectines de type C (C-type lectins and phagocytosis 2009), comme :
- la dectine-1 (CLEC7A),
- la dectine-2 (CLEC6A),
- Mincle (Macrophage inducible Ca2+-dependent lectin receptor) ou
- DC-SIGN (Dendritic Cell-Specific ICAM3- Grabbing Non-integrin, aussi appelée CD209) ;
2. Certaines molécules de reconnaissance sont de type lectine tel que CD33.
Remarque : CD signifie Cluster of Differenciation ou classes de différenciation, i.e. glycoprotéines membranaires classées selon une nomenclature utilisée pour l'identification et l'immunophénotypage des cellules du système immunitaire, et jouant un rôle de marqueurs de coanticorps.
3. Les récepteurs scavenger, i.e. éboueurs en français, reconnaissent et dégradent les LDL (lipoprotéines de basse densité) modifiées par acétylation.

Les récepteurs scavenger sont étudiés dans des chapitres spéciaux.
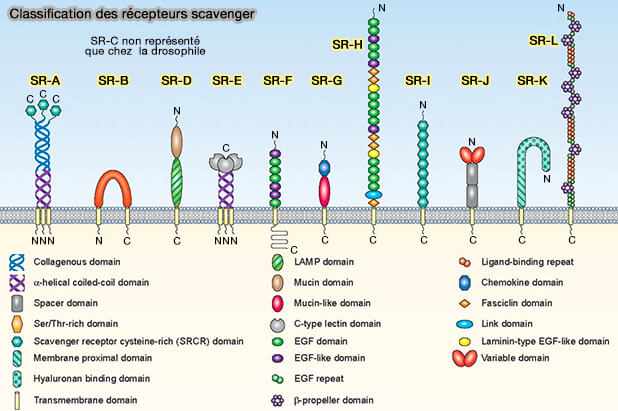
(Figure : vetopsy.fr d'après PrabhuDas et coll)
Remarque : bien que les récepteurs Toll-like (TLR) puissent également détecter des modèles moléculaires sur les agents pathogènes, ils ne sont pas des récepteurs phagocytaires.
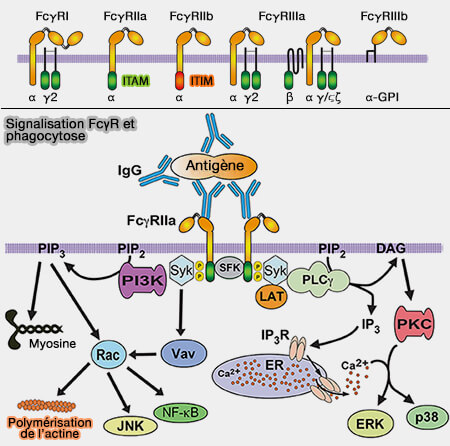
(Figure : vetopsy.fr d'après Uribe-Querol et Rosales)
Néanmoins, les TLR peuvent coopérer avec les récepteurs phagocytaires pour rendre la phagocytose plus efficace (Control of adaptive immunity by the innate immune system 2015).
Récepteurs opsoniques
Les récepteurs opsoniques détectent les opsonines, i.e. protéines dérivées de l'hôte, et marquent les particules comme cibles de la phagocytose.
1. Les anticorps, i.e. les FcR, i.e. Fragment crystallisable Receptor, reconnaissent le fragment FC des immunoglobulines G (IgG) ou IgA (Fc receptors: Cell activators of antibody functions 2013).
2. La fibronectine qui joue un rôle essentiel dans l'adhérence entre cellule et matrice extracellulaire (MEC).
3. Le complément forme un groupe d'environ 50 protéines connues du sérum, faisant partie de l'immunité innée, i.e. les récepteurs du complément se lient aux composants activés du complément, tels que iC3b, déposés sur la particule (Macrophage complement receptors and pathogen clearance 2007).
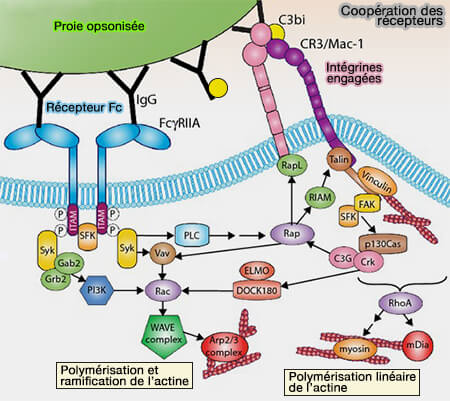
(Figure : vetopsy.fr d'après Freeman et Grinstein)
4. La globuline de graisse du lait (lactadhérine ou MFGE8) peut fonctionner comme une protéine d'adhésion cellulaire pour connecter le muscle lisse aux fibres élastiques dans les artères.
5. La lectine liant le mannose (MBP) active le système du complément du système immunitaire par la voie des lectines, afin de former un complexe d'attaque membranaire (MAC).
Remarque : Pour une reconnaissance efficace de la particule cible et l'initiation de la phagocytose, de nombreux récepteurs sur la membrane phagocytaire doivent interagir avec plusieurs molécules d'IgG sur la particule opsonisée.
Contacts récepteurs/ligands
Problématique
1. Les récepteurs membranaires, dans leur grande majorité, à part quelques récepteurs scavengers, sont des molécules courtes qui dépassent de 5 nm seulement le feuillet externe de la bicouche.
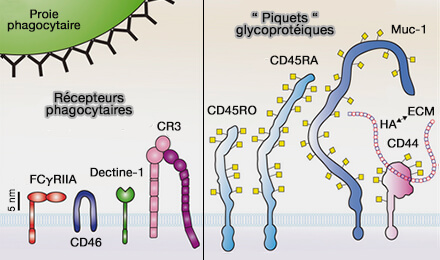
des piquets glycoprotéiques
(Figure : vetopsy.fr d'après Ostrowski et coll)
2. Par contre, les glycoprotéines transmembranaires, majoritaires dans la membrane et constituant une partie du glycocalyx, sont beaucoup plus longues et rigides, i.e. celles des leucocytes vont réduire l'accès du ligand aux récepteurs courts.
a. C'est le cas des phosphatases transmembranaires :
- CD45/PTPRC), i.e. Protein tyrosine phosphatase, receptor type C,
- CD148/PTPRJ, i.e. Receptor-type tyrosine-protein phosphatase eta,
b. Les mucines transmembranaires comme Muc-1 sont des protéines hautement glycosylées par les liaisons O-GalNAc au niveau des résidus sérine et thréonine ( O-glycosylation).
O-glycosylation).
c. CD44 et les hyaluronanes génèrent la matrice péricellulaire.
Par exemple, la CD45 des lymphocytes T est trois fois plus grande que les récepteurs des cellules T (TCR) et érige une barrière perpendiculaire à la membrane plasmique pour l'engagement des récepteurs phagocytaires courts (Initiation of T cell signaling by CD45 segregation at ‘close-contacts’ 2016).
Hypothèses
S'il existe une couche haute et dense de glycocalyx autour de la cellule, comment des récepteurs courts contactent-ils leurs ligands ? Trois hypothèses peuvent être envisagées.
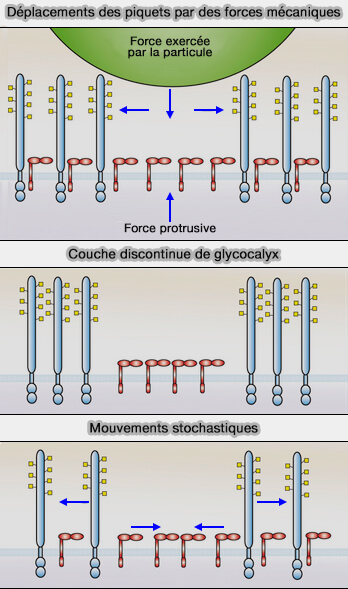
(Figure : vetopsy.fr d'après Ostrowski et coll)
1. La plus probable est que la force mécanique exercée par la particule approchante et/ou par le phagocyte lui-même, lorsqu'il étend activement ses protrusions, peut suffire à déplacer le glycocalyx latéralement, amenant les ligands en apposition étroite avec les récepteurs.
En effet, l'application d'une force exogène sur les cellules ou l'augmentation de la rigidité des substrats sur lesquels elles sont cultivées conduit au déplacement de grandes glycoprotéines des contacts adhésifs (The cancer glycocalyx mechanically primes integrin-mediated growth and survival 2014).
2. La couche de glycocalyx peut être discontinue et des zones peuvent ainsi exposer leurs récepteurs courts, ce qui semble être le cas dans les cellules endothéliales (Salt overload damages the glycocalyx sodium barrier of vascular endothelium 2011).
Toutefois, les régions de la membrane des macrophages qui entrent en contact en premier avec les particules cibles sont, en fait, richement dotées en CD44 et CD45.
3. Le mouvement stochastique des molécules les plus hautes peut momentanément exposer les récepteurs les plus courts, leur permettant d'entrer en contact avec la particule cible, ce qui est peu probable, car des zones exposées relativement grandes devraient être générées pour permettre l'accès à des particules rigides de grande taille.

Les trois hypothèses précédentes ne s'excluent pas mutuellement et nécessitent des tests expérimentaux plus poussés.
Diffusion membranaire des glycoprotéines
La ségrégation du glycocalyx des récepteurs pour initier la signalisation et le regroupement ultérieur des récepteurs phagocytaires nécessite une diffusion latérale des glycoprotéines et des récepteurs dans le plan de la membrane plasmique.
1. En effet, de nombreuses glycoprotéines agissent comme des " piquets " pour les " clôtures " cytosquelettiques car elles possèdent :
- des domaines cytoplasmiques qui se lient directement au cytosquelette sous-membranaire, i.e. au cortex cellulaire,
- des ectodomaines se lient souvent aux matrices péricellulaires ou extracellulaires.
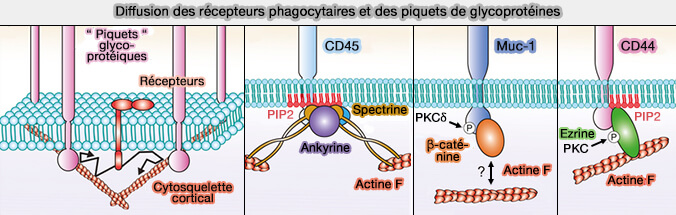
(Figure : vetopsy.fr d'après Ostrowski et coll)
2. Ces piquets constituent des obstacles (relativement) immobiles et en même temps un pont entre les polymères cytosquelettiques et péricellulaires qui restreignent la diffusion d'autres molécules membranaires plus mobiles.
Le déplacement de grosses glycoprotéines atténue donc deux obstacles à l'engagement des récepteurs :
- la barrière stérique perpendiculaire à la membrane,
- les contraintes à la diffusion latérale.
Retour à la phagocytose
Biologie cellulaire et moléculaireConstituants de la celluleTransport membranaireTransports sans mouvements membranairesTransports passifsTransports actifsTransports avec mouvements membranaires : trafic vésiculaireFusion/fission membranaireEndocytoseEndocytose clathrine-dépendante (CME)Endocytoses clathrine-indépendantes (CIE)CavéolesFEMEVoie CLIC/GEECVoie flotillineVoie Arf6IL2RGFR-NCEADBEUFEPinocytoseMacropinocytosePhagocytoseVoie sécrétoireCanaux ioniquesTransporteursUniportsPompesCo-transporteursMoteurs moléculairesVoies de signalisation