Trafic vésiculaire
Endocytose clathrine-indépendante (CIE)
Endocytose rapide endophiline-dépendante (FEME)
Recrutement de l'endophiline, amorçage, sélection des cargos et courbure membranaire
L'endocytose rapide endophiline-dépendante (FEME) est une endocytose indépendante de la clathrine (CIE), régulée par l'endophiline, qui permet le recrutement et l'amoçage pour le transport de cargos spécifiques.
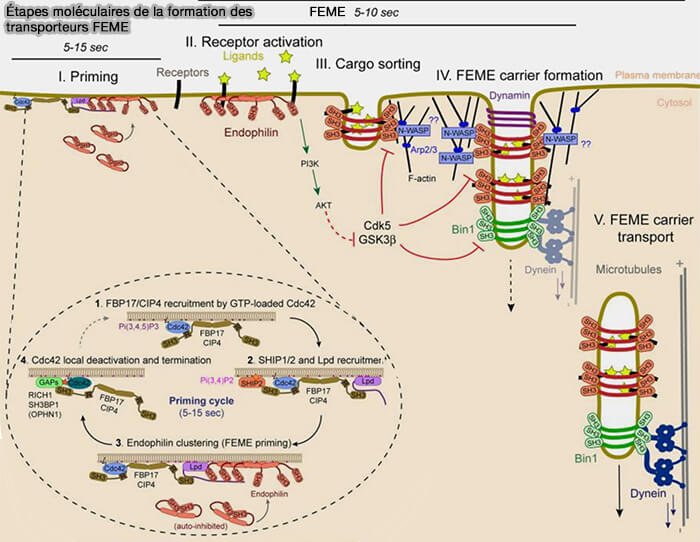
Étapes moléculaires de la formation des transporteurs FEME
(Figure : vetopsy.fr d'après Sacramento et Boucrot)
Recrutement de l'endophiline
Dans les cellules au repos, l'endophiline est préenrichie en petits patchs à des emplacements discrets sur la membrane plasmique.
L'engagement du domaine SH3 avec un motif riche en proline (PRM ou PRD) est nécessaire et suffisant pour lever l'auto-inhibition et pour que le domaine N-BAR puisse se lier à la membrane pour détecter, stabiliser ou induire une courbure membranaire locale.
2. Les amas d'endophiline sur la membrane plasmique dépendent de la production de Pi(3,4)P2 à partir de précurseurs Pi(4,5)P2 et Pi(3,4,5)P3.
Ils sont donc imposants au bord d'attaque des cellules migratrices, mais sont également nombreux sur les surfaces ventrales et dorsales des cellules.
3. Les amas d'endophiline sur la membrane plasmique des cellules au repos sont dynamiques et durent de 5 à 15 s avant leur désassemblage si l'endocytose n'a pas lieu.
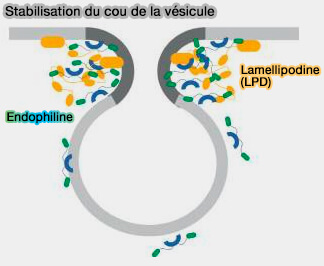
Stabilisation du cou vésiculaire
(Figure : vetopsy.fr d'après Mondal et coll)
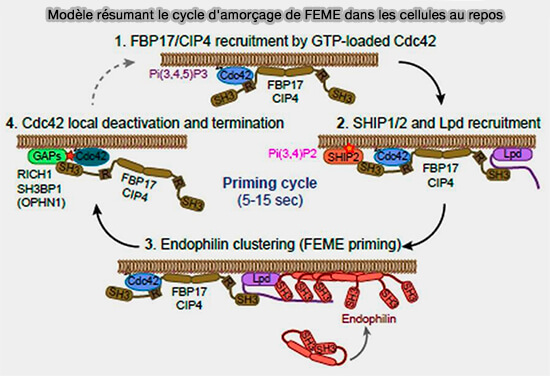
Dans certaines cellules, TOCA1 (Transducer of Cdc42-dependent actin assembly protein 1) pourrait être impliquée dans cette étape, car elle s'hétérodimérise à la fois avec FBP17 et CIP4, et tous les trois sont fonctionnellement redondants dans la FEME (Investigation of the Interaction between Cdc42 and Its Effector TOCA1 2016).
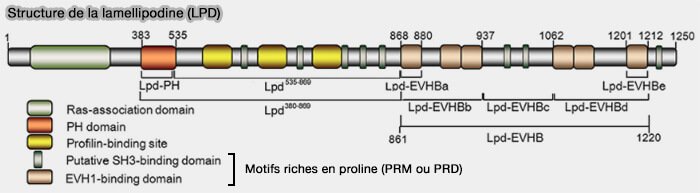
2. CIP4 (Cdc42-Interacting Protein4) et FBP17 recrutent, via leur domaine SH3, SHiP1/2 (SH2-containing inositol Phosphatase 1/2) et la lamellipodine (LPD).
3. L'endophiline A se lie aux régions de la LPD, concentrant ainsi l'endophiline en plaques dans la membrane plasmique.
Modèle résumant le cycle d'amorçage de FEME dans les cellules au repos
(Figure : vetopsy.fr d'après Chan Wah Hak et coll)
Ces trois événements sont des étapes obligatoires qui doivent se produire de manière séquentielle.
L'épuisement de l'une quelconque de ces protéines délimite le processus en aval de l'étape visée, qui bloque invariablement la formation de vecteur FEME lors de l'activation du récepteur.
4. En l'absence d'activation des récepteurs, les patchs d'endophiline se désassemblent rapidement en 5 à 15 secondes après le recrutement local des GAP (GTPase-Activating Protein ou GTPase-Accelerating Protein) de Cdc42, Rich1, Sh3BP1 ou Oligophrénine-6.
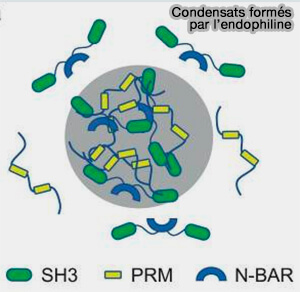
Condensats formés par l'endophiline
(Figure : vetopsy.fr d'après Mondal et coll)
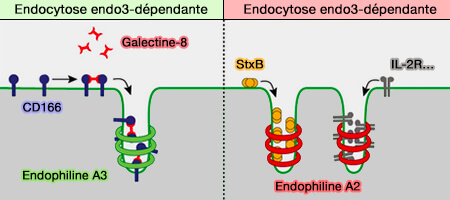
Endocytoses A3 et A2-dépendantes
(Figure : vetopsy.fr d'après Renard et coll)
Remarque : cependant, de nombreux cargos peuvent également utiliser d'autres voies endocytaires pour entrer dans les cellules, par exemple par CME, CLIC/GEEC, EGFR-NCE ou macropinocytose.
2. Lors de la formation rapide de transporteurs FEME, i.e. l'endophiline peut se lier :
2. Le domaine SH3 de l'endophiline se lie à ces motifs PRM.
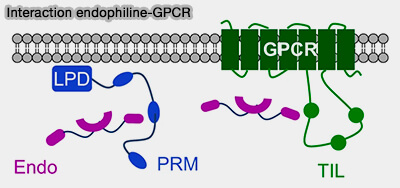
Les dimères d'endophiline relient directement la membrane par l'intermédiaire de leurs domaines BAR aux effecteurs cytosoliques et aux cargos par l'intermédiaire de leurs domaines SH3.
Récepteurs à activité tyrosine kinase (RTK)
L'endophiline se lie indirectement aux récepteurs à activité tyrosine kinase (RTK) par l'intermédiaire d'une protéine adaptatrice reconnaissant des motifs dans leurs queues cytoplasmiques.
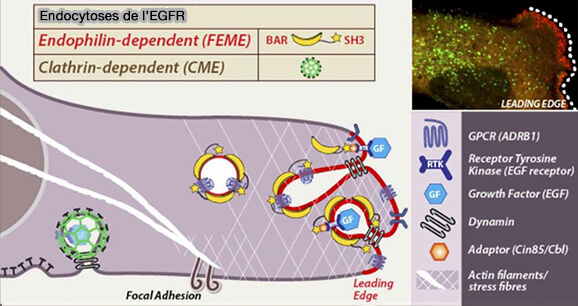
1. Dans le cas de l'EGFR (Epidermal Growth Factor Receptor) et HGFR (Hepatocyte Growth Factor Receptor), seuls les récepteurs activés, i.e. dimérisés et transphosphorylés, sont reconnus par Cbl par son domaine SH2, qui est à son tour lié par les complexes CIN85-Endophiline.
L’internalisation de l’EGFR dépend de la concentration d’EGF.
À forte concentration, le récepteur est internalisé pour la destruction lysosomale, en plus de la CME, par la FEME ou par la voie EGFR-NCE (Non Clathrin-Endocytosis) qui est une voie alternative.
Cbl se lie aux tyrosines phosphorylées et est, en retour, phosphorylée par le récepteur.
Cette phosphorylation permet le recrutement de la protéine adaptatrice CIN85/SH3KBP1 qui est associée aux endophilines, et le complexe CIN85/EGFR/endophilines ainsi formé entraine l’endocytose du récepteur stimulé.
Endocytoses de l'EGFR
(Figure : vetopsy.fr d'après Boucrot et coll)
Remarque : EGFR peut aussi être endocyté par la voie EGFR-NCE (Non Clathrin-Endocytosis), qui est activée à concentrations élevées et cible les récepteurs de la dégradation, les lysosomes, atténuant la signalisation.
Le processus de courbure de la membrane dans la FEME n'est pas encore complètement identifié.
1. L'endophiline et son domaine N-BAR sont nécessaires, car en leur absence, aucun transporteur FEME n'est produit.
2. Cependant, la transition entre les plaques primitives de l'endophiline, qui ne semblent pas générer de courbure significative, et la mise en forme des tubules et des vésicules après l'activation des récepteurs ne sont pas comprises, i.e. les patchs d'endophiline dans les cellules au repos ne correspondent pas aux invaginations membranaires.
Soit les patchs primitifs n'atteignent pas une concentration locale critique d'endophiline nécessaire pour déformer la membrane plasmique, et l'activation du récepteur augmente le nombre de molécules d'endophiline au-dessus du seuil.
Soit, jusqu'à présent, un mécanisme non identifié contrôle la courbure de la membrane par le domaine N-BAR jusqu'à l'activation.
Soit la courbure nécessite la présence d'une autre protéine.
Bin1 est une autre protéine N-BAR présente sur pratiquement tous les transporteurs FEME, mais aussi sur la plupart des patchs d'amorçage, de sorte que son recrutement n'est pas le déclencheur en soi ( régulation de la FEME).
3. La polymérisation de l'actine pourrait avoir un rôle dans l'induction ou la stabilisation de la courbure de la membrane au cours de la formation des tubules.
Mais isoler son rôle lors de l'amorçage (priming), la formation de courbure de la membrane, la scission de membrane et le transport cytosolique potentiel à courte distance est techniquement difficile.





 régulation de la FEME),
régulation de la FEME),