


Constituants cellulaires
Cytosquelette : microtubules
Fission : mécanisme
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Matrice extracellulaire
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Les AAA+ exercent une traction séquentielle sur l’E-hook de la tubuline, utilisant des cycles ATP pour extraire un dimère et provoquer la coupure microtubulaire.
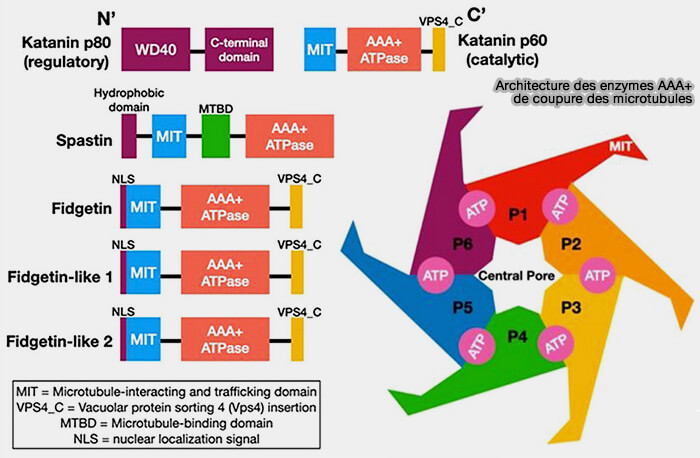
(Figure : vetopsy.fr d'après Smart et Sharp)
Fonctionnement général des AAA+ de coupure des MT
Avant la coupure
Les sous-unités existent en solution sous forme de monomères ou d’oligomères partiels.
1. L'enzyme s'ancre sur le microtubule par un domaine spécialisé :
- fidgetine via des déterminants faibles et diffus, patchs basiques, signaux locaux de tyrosination, et interactions avec des partenaires, plutôt que sur un domaine spécialisé de type MTBD ou MIT.
- la spastine, via le domaine MTBD,
- la katanine via sa sous-unité p80.
2. Les sous-unités AAA+ s’associent progressivement, formant une spirale ouverte (pré-annea), hexamère qui ne devient n’est stable que lorsqu’il reconnaît un substrat, ici l'E-hook de la tubuline
Pendant la coupure
Les enzymes AAA+ impliquées dans la fission microtubulaire (katanine, spastine et fidgetine) partagent un mode d’action mécanique commun fondé sur un hexamère en anneau utilisant l’énergie de l’ATP pour extraire des dimères de tubuline et rompre le lattice (Katanin spiral and ring structures shed light on power stroke for microtubule severing 2020).
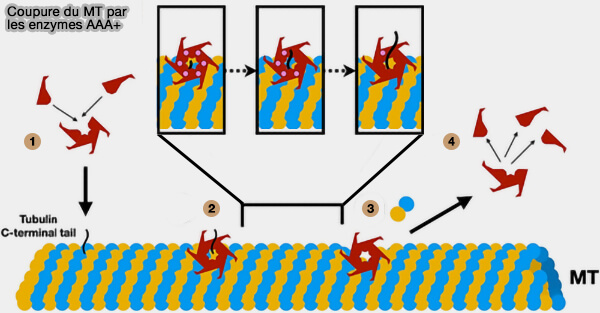
(Figure : vetopsy.fr d'après Smart et Sharp)
1. Chaque enzyme s’organise en un anneau hexamérique, dont le centre forme un pore.
a. L’hexamère AAA+ est représenté sous deux formes :
- spiralée, asymétrique, en état de pré-engagement
- fermée en anneau, symétrique, état actif.
b. Le passage de l’une à l’autre est couplé à la liaison/hydrolyse de l’ATP et prépare la séquence de traction, i.e. powerstroke.
2. Chaque sous-unité AAA+ possède une pore loop dirigé vers le centre de l’anneau qui interagit avec le C-terminal de la tubuline (E-hook).
a. La pore loop se déplace verticalement et se rapproche du pore lors de la transition spirale/anneau.
b. Le segment flexible du C-terminal de la tubuline (E-hook) i dépend du type d’ATPase :
- La fidgetine intervient sur E-hooks peu modifiés (tyrosinés).
- La spastine intervient sur E-hooks modérés, sensibles aux modifications post-traductionnelles (PTM),
- La katamine intervient E-hooks fortement polyglutamylés.
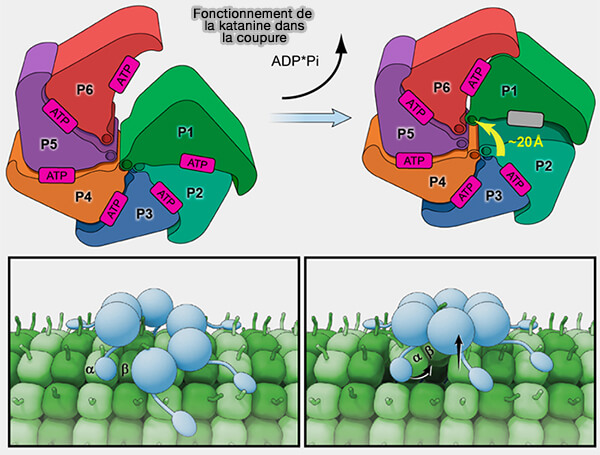
(Figure : vetopsy.fr d'après Zehr et coll)
3. La dégradation séquentielle de l’ATP par les sous-unités génère une traction par à-coups sur l’E-hook, engagée par les pore loops du domaine AAA+, i.e. power stroke.
- L’E-hook reste engagé dans le pore après le premier power stroke.
- À chaque nouvelle hydrolyse, la sous-unité suivante abaisse son pore loop dans l’axe de l’anneau, transloquant la tubuline un peu plus vers l’intérieur du pore et l’extrayant progressivement de son lattice.
4. Sous cette tension croissante, on asssite à une déformation et une rupture du lattice, i.e. les contacts latéraux entre protofilaments cèdent, les contacts longitudinaux se rompent, le protofilament se désolidarise localement.
- Lorsque la contrainte dépasse le seuil mécanique du lattice, un dimère α/β-tubuline est extrait, créant une discontinuité.
- La rupture se propage alors en une coupure franche du microtubule.
Remarque : le mécanisme est identique dans son principe pour les trois enzymes, mais diffère par :
- la force générée, i.e. fidgetin < spastine < katanine,
- la nature des microtubules ciblés, jeunes vs modifiés vs très polyglutamylés,
- le contexte cellulaire, i.e. migration, neurites, centrosome, fuseau.
Apès la coupure
1. Après la coupure, le dimère extrait, qui comprend deux extrémités nouvellement exposées, une (+) et une (-), peut (Cutting, Amplifying, and Aligning Microtubules with Severing Enzymes 2021) :
- se dépolymériser, ce qui correspond à une catastrophe locale initiée par l’absence de coiffe GTP sur l’extrémité nouvellement formée,
- se sauver si des GTP-islands sont proches, car la rencontre d’un patch GTP-like suffit à redresser les protofilaments et à rétablir une géométrie propice à la repolymérisation ;
- servir de matrice pour de nouveaux microtubules, puisque les fragments ainsi générés constituent des noyaux structuraux capables de réamorcer une croissance rapide dans les régions où le réseau doit être densifié ou réorganisé.
Remarque : le dimère peut aussi être dirigé vers le protéasome s''il est dénaturé lors de stress par exemple.

(Figure : vetopsy.fr d'après Kuo et Howard)
2. Une fois la traction terminée, l’hexamère ne s’effondre pas totalement, mais perd sa configuration serrée.
- Certaines sous-unités se détachent ou se relâchent.
D’autres restent temporairement associées au microtubule.
L’enzyme peut être recyclée ou relancer un cycle si un autre E-hook est proche.
Retour à la fission des microtubules
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosqueletteFilaments d'actineFilaments intermédiairesMicrotubulesTubulineDynamique des MicrotubulesNucléationFissionMAPCentrioleCentrosomeFuseau mitotiqueReproduction cellulaireBiochimieTransport membranaire Moteurs moléculairesVoies de signalisation