L'apolipoprotéine E (apoE) est une protéine plasmatique impliquée dans le métabolisme des lipides et, en tant que composant des lipoprotéines, médie leur absorption cellulaire en se liant aux récepteurs de surface cellulaire.
Deux sites (Thr-26 et Thr-36) sont dans la région N-terminale (région de liaison au récepteur),
deux (Thr-212 et Ser-215) dans la région charnière ou dans l'extrémité C-terminale selon les auteurs, i.e. la numérotation des résidus n'étant pas identique, ,
trois sites (Thr-307, Ser-308 et Ser-314) à l'extrémité C-terminale (région de liaison aux lipides).
On ne peut extrapoler les rôles de l'apoE dans le cerveau aux tissus périphériques.
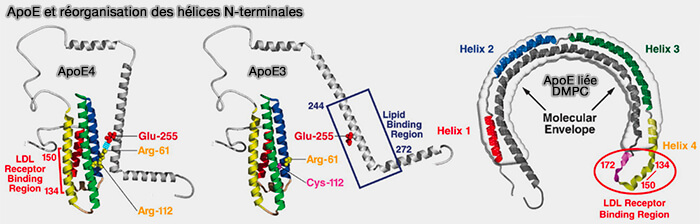
Sa structure comprend des ponts salins inter et intra-hélicoïdaux et des interactions hydrophobes telles que l'empilement aromatique et les glissières à leucine (leucine zipper) conférant une résistance au dépliement et à l'ouverture du faisceau d'hélice.
Les interactions hélice/hélice sont plus fortes que celles hélice/lipide, une caractéristique essentielle à son rôle de ligand pour LDLR, i.e. le faisceau d'hélice, empêchant ainsi une clairance prématurée.
C1 et C2 seraient pliées en hélices α amphipathiques de classe A, i.e. résidus chargés positivement regroupés à l'interface polaire/non polaire et résidus chargés négativement au centre de la face polaire, préférant les interactions hélice-lipide aux interactions hélice-hélice.
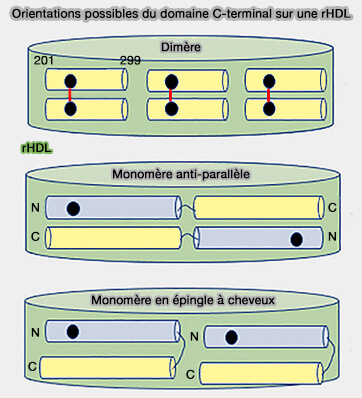
Orientations du domaine C-terminal sur une rHDL
(Figure : vetopsy.fr d'après Kothari et coll)
2. Ainsi, le domaine C-terminal ancre la protéine entière à une particule de lipoprotéine, permettant au domaine N-terminal d'être :
soit dans un état de faisceau d'hélice sans lipide,
soit dans un état ouvert lié aux lipides.
Les modèles de cette interaction avec les lipoprotéines peuvent prendre plusieurs orientations dans les rHDH, i.e. HDL résiduelles ou remnants de HDL :
anti-parallèle ou en épingle à cheveux pour les monomères,
parallèle pour les dimères.
3. La transition entre les conformations ouvertes et en faisceau d'hélice d'une apoE ancrée semble être régulée par la disponibilité des surfaces lipidiques hydrophobes sur une particule de lipoprotéine ( pression de surface).
L'ancrage de l'apoE à une surface lipidique est une première étape essentielle pour faciliter l'ouverture du faisceau d'hélices et l'interaction des récepteurs du domaine N-terminal, conduisant à une éventuelle clairance cellulaire de la particule de lipoprotéine du plasma.
1. La première étape, réversible et rapide, implique l'interruption induite par la liaison des lipides des interactions de domaine, favorisant la dissociation séquentielle du domaine C-terminal et des domaines charnières du domaine N-terminal.
2. La deuxième étape, réversible et lente, implique l'ouverture du faisceau d'hélices NT à l'extrémité de la boucle.
La structure finale associée aux lipoprotéines de l'apoE3 est une conformation complètement ouverte, contenant deux lobes C et N-terminal reliés par le domaine charnière
Les deux lobes interagissent avec la surface des lipoprotéines, fournissant la surface de liaison aux lipoprotéines maximale possible.
La principale région de liaison au LDLR dans cette conformation est entièrement exposée en raison de la suppression des interactions entre les hélices 3 et 4 ce qui entraîne un potentiel de charge positive accru pour l'activité de liaison au récepteur optimisée.
3. Ce processus est essentiel car il garantit que seule l'apoE entièrement lipidée est capable de se lier aux LDLR avec une activité optimale
La principale région de liaison au LDLR est protégée par le domaine C-terminal qui empêche l'apoE sans lipides de se lier prématurément aux récepteurs de l'apoE.
La conformation inactive du faisceau d'hélices N-terminal sans lipides empêche à la fois l'apoE sans lipides et partiellement lipidée de se lier au récepteur.
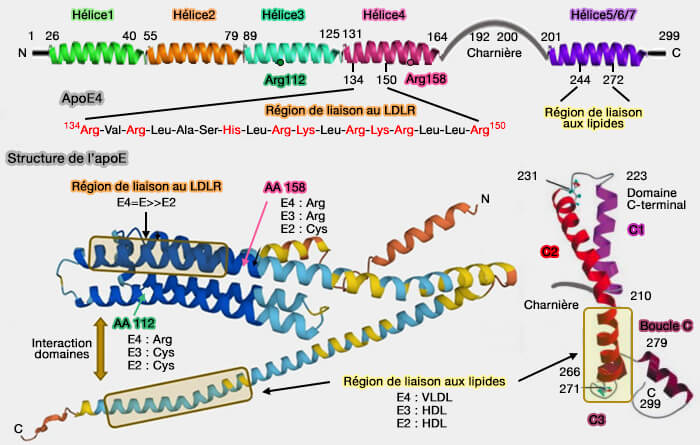
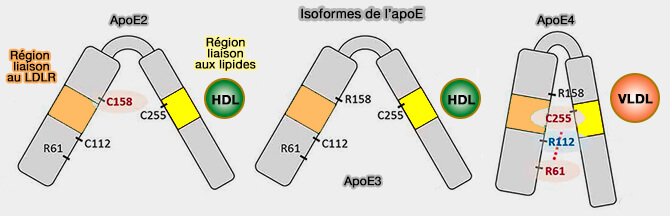
L'apoE2 contient un résidu cystéine à cette position qui supprime un pont salin entre Arg158 et Asp154, ce qui entraîne un changement conformationnel du domaine de liaison au récepteur et finalement une liaison défectueuse au LDLR (<2 % de l'activité de liaison normale au LDLR) et au LRP1, i.e. 30 à 50 % de l'activité normale de liaison au LRP1 (Salt bridge relay triggers defective LDL receptor binding by a mutant apolipoprotein 1994).
a. L'interaction entre les acides aminés 61 et 112 influence la liaison aux lipoprotéines.
C'est la principale raison expliquant la forte affinité de l'apoE4 pour les VLDL tandis que l'apoE3 et l'apoE2 se lient aux HDL (lipoprotéines de haute densité).
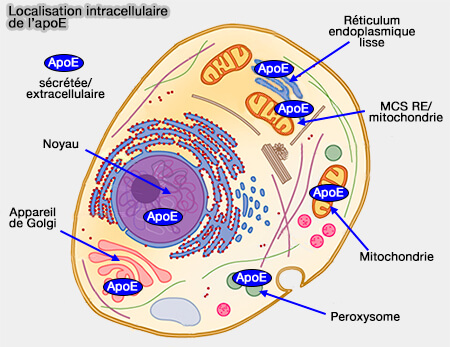
La sécrétion de cargos mal repliés par le réticulum endoplasmique (RE) et structurellement modifiés dans les mitochondries peut expliquer l'apparition de fragments apoE dans la mitochondrie.
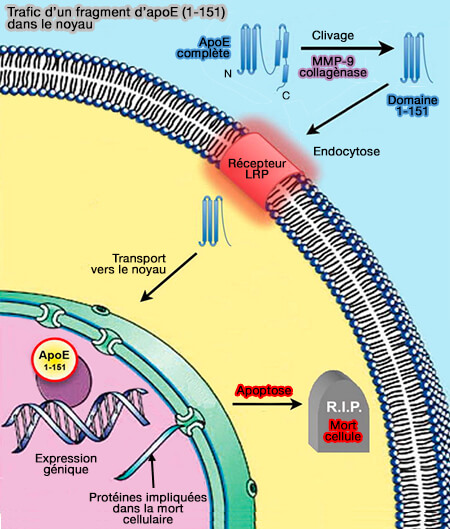
Le clivage protéolytique de l'apoE donne des fragments avec des caractéristiques de repliement altérées, qui peuvent être reconnus comme mal repliés et donc transférés aux mitochondries pour la séquestration.
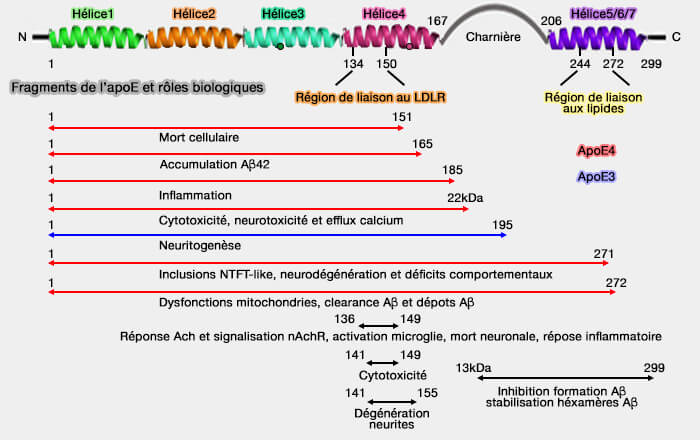
Protéolyse et fragments d'apoE
Divers fragments d'apoE, de longueur variable (quelques acides aminés jusqu'à 29 kDa) sont généralement tronqués à l'extrémité C-terminale, résultant de clivage protéolytique sont retrouvés.
Les fragments apoE détectés ont des effets indésirables.
Fragments de l'apoE et rôles biologiques
(Figure : vetopsy.fr d'après Munoz et coll)
une altération de la fonction mitochondriale en se liant directement aux protéines de la chaîne respiratoire mitochondriale et diminue l'activité des complexes III et IV,






 apolipoprotéines et groupes de gènes et Functional diversity of apolipoprotein E: from subcellular localization to mitochondrial function 2022).
apolipoprotéines et groupes de gènes et Functional diversity of apolipoprotein E: from subcellular localization to mitochondrial function 2022).