


Enzymes
Groupe des
transférases (EC 2)
Protéine kinases
Sérine/thréonine kinases
CaMII kinases
(CaMKII) : structure et rôles
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Enzymes
- Vue d'ensemble
- Classification des enzymes
- Oxydoréductases (EC 1)
- Transférases (EC 2)
- Hydrolases (EC 3)
- Lyases ou synthases (EC4)
- Isomérases (EC5)
- Ligases ou synthétases (EC 6)
- Coenzymes
- Vitamines
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La CaMKII couple une organisation structurale en anneau à des mécanismes d’autoactivation, permettant l’intégration temporelle des signaux calciques et la régulation durable de multiples processus cellulaires.
Structure de la CaMKII
1. La CaMKII s'assemble en une structure dodécamérique (12 sous-unités) et regroupe pas moins de 28 isoformes, les α et β prévalent dans le cerveau.
Dans l'holoenzyme, les domaines catalytiques N-terminaux sont dirigés vers l'extérieur, ce qui leur permet de phosphoryler des résidus entre les sous-unités.
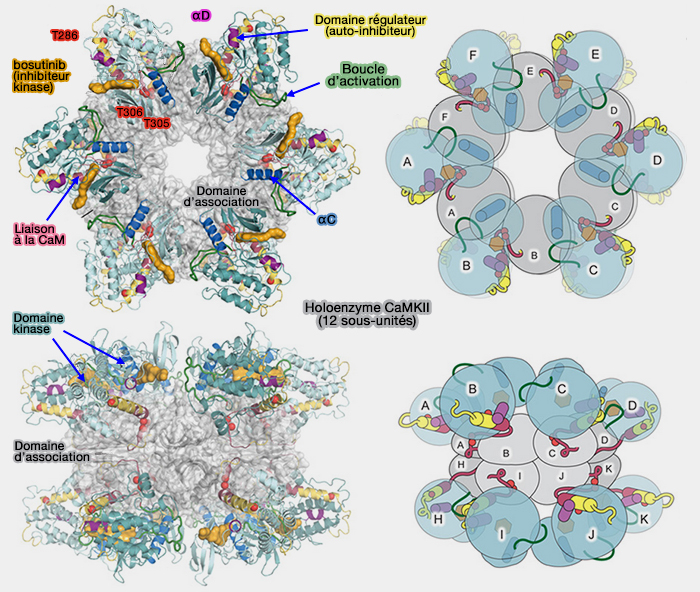
(Figure : vetopsy.fr d'après Chao et Stratton)
2. Les CaM kinases sont formées par (A mechanism for tunable autoinhibition in the structure of a human Ca2+/calmodulin-dependent kinase II holoenzyme 2011) :
- un domaine catalytique N-terminal,
- un domaine auto-inhibiteur ou régulateur,
- un domaine variable,
- un domaine d'association.
-
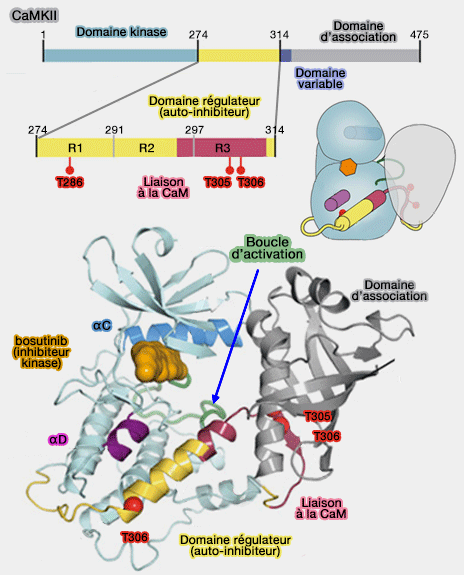
Structure de la CaMKII
(Figure : vetopsy.fr d'après Chao et Stratton)
Domaine catalytique
Le domaine catalytique a plusieurs sites de liaison pour l'ATP et d'autres protéines d'ancrage du substrat.
Il est responsable du transfert du phosphate de l'ATP vers les résidus Ser ou Thr dans les substrats.
Domaine auto-inhibiteur
1. Le domaine auto-inhibiteur (ou régulateur) est classiquement divisé en 3 régions (R1, R2 et R3).
- Il se lie au domaine catalytique et bloque sa capacité à phosphoryler les protéines.
- Il est déplacé par la liaison du module Ca++/calmoduline.
2. La caractéristique structurelle qui régit cette auto-inhibition est le résidu T286 chez la souris à la base d'une hélice α formée par le segment régulateur.
- Cette hélice α bloque la liaison du substrat dans la structure auto-inhibée en occupant un sillon hydrophobe sur le domaine kinase.
- Sa phosphorylation se produit en trans, entre deux sous-unités de la kinase de la même holoenzyme active de manière permanente CaMKII.
Les deux autres sites phosphorylables sont les T305 et T306 du domaine R3 qui se trouvent au centre du site de liaison à la CaM.
Domaine variable
Le domaine variable, lien flexible entre les domaines précédant et suivant, porte bien son nom et dépend des différentes isoformes de CaMKII.
Domaines d'auto-association
(Figure : vetopsy.fr d'après Chao et Straton)
Le domaine d'auto-association, localisé à l'extrémité C-terminale, permet l'assemblage des protéines individuelles en grands multimères (12 sous-unités) de deux cycles de 6 sous-unités l'un sur l'autre (Structure of the CaMKIIδ/Calmodulin Complex Reveals the Molecular Mechanism of CaMKII Kinase Activation 2010).
Fonctionnement
de la CaMKII
La CaMKII est inactive dans les conditions de repos (CaMKII regulation in information processing and storage 2012 et Structural studies on the regulation of Ca2+/calmodulin dependent protein kinase II 2013).
Vue d'ensemble
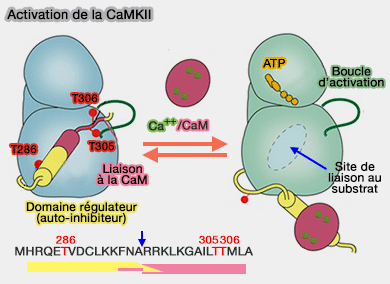
1. En l'absence de Ca++, l'accès du substrat au site de liaison du domaine catalytique est bloqué par le domaine auto-inhibiteur de la CaMKII.
2. Lors de l'afflux de Ca++, la liaison de Ca++ à la calmoduline conduit au complexe Ca++/CaM qui se lie à deux sous-unités voisines et conduit à l'autophosphorylation de T286.
(Figure : vetopsy.fr d'après Straton et Chao)
La phosphorylation de T286 perturbe la position des éléments R1 et R2 du segment de régulation contre le domaine kinase et lève l'auto-inhibition.

La phosphorylation de T286 confère une activité indépendante du calcium à la kinase et permet une activation permanente de l'enzyme CaMKII, même après l'élimination de Ca++/CaM.
a. En effet, les sous-unités CaMKII activées s'autophosphorylent les unes les autres.
- Si une sous-unité T286 est déphosphorylée, elle se rephosphoryle lorsque la sous-unité voisine est encore phosphorylée.
b. Cette autophosphorylation est renforcée par la structure de l'holoenzyme qui se présente sous la forme de deux anneaux empilés (CaMKII Autonomy Is Substrate-dependent and Further Stimulated by Ca2+/Calmodulin 2010).
- La phosphorylation augmente considérablement l'affinité pour le complexe calmoduline, processus appelé piégeage de la CaM, ce qui prolonge la durée d'activité de la kinase.
- L'autophosphorylation des 12 sous-unités au sein d'une holoenzyme permet une translation graduelle de la fréquence des pointes de Ca++ dans l'activité kinase (Nonlinear Decoding and Asymmetric Representation of Neuronal Input Information by CaMKIIα and Calcineurin 2013).
c. En outre, l'inhibition de la PP1 (protéine phosphatase I) favorise aussi l'autophosphorylation et permet à CaMKII d'être constamment actif.
3. La déphosphorylation de T286 provoque l'inactivation de CaMKII.
La phosphorylation de T305 et T306 inhibe la fixation de la calmoduline.
Changements conformationnels
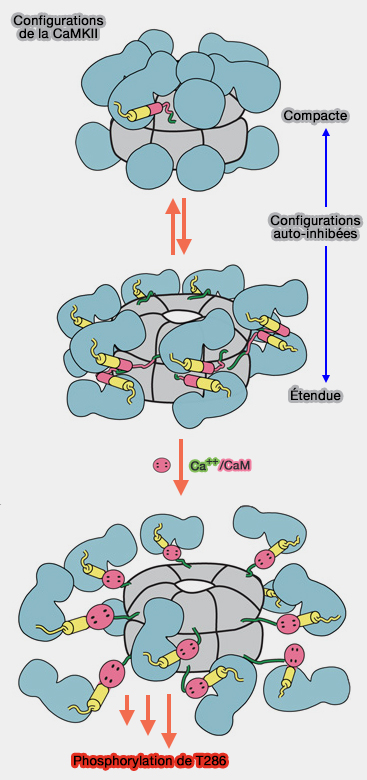
1. L'holoenzyme CaMKII, à l'état auto-inhibé, présente deux configurations.
- La première configuration est compacte : le domaine de liaison à la calmoduline interagit avec le centre de l'holoenzyme, ce qui le rend inaccessible à la calmoduline.
- L'autre configuration est étendue : le domaine de liaison à la calmoduline est plus exposé, sans que cette structure ait été explicitée pour l'instant.
Un équilibre existe entre ces états, et est sensible à la longueur du lien variable reliant les domaines catalytique, auto-inhibiteur et d'association.
(Figure : vetopsy.fr d'après Anderson)
2. Lors de la liaison Ca++/ CaM, le segment régulateur est libéré du domaine kinase, ce qui ouvre complètement la structure (New Therapeutic Targets in Cardiology: Arrhythmias and CaMKII 2013)
.Implications
de la CaMKII
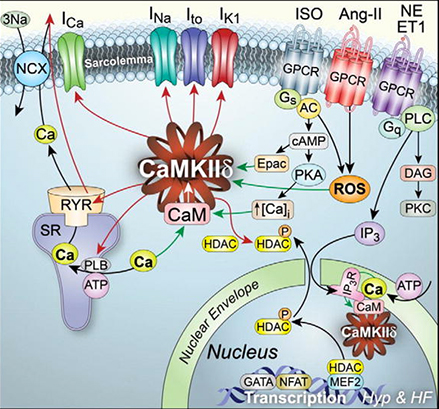
La CaMKII est impliquée dans la phosphorylation de nombreuses protéines.
1. CaMKII interagit avec des canaux ioniques :
- les canaux calciques voltage-dépendants (Cav), par les sous-unités α1 et β (
 régulations des canaux Cav par les kinases),
régulations des canaux Cav par les kinases), - les canaux sodiques voltage-dépendants (Nav),
- les canaux potassiques voltage-dépendants.
2. CaMKII interagit avec les gènes.
3. CaMKII interagit avec de nombreuses protéines de la matrice extracellulaire (MEC) actives dans :
- l'organisation post-synaptique (CaMKII: Claiming Center Stage in Postsynaptic Function and Organization 2014)
- les cancers (The emerging role of CaMKII in cancer 2015).
BiochimieChimie organiqueBioénergétiqueProtidesGlucidesLipidesEnzymesKinasesProtéines kinasesLipides kinasesHydrolasesPeptidases/protéasesPhosphatasesEstérasesLipasesPhospholipasesATPasesGTPasesProtéines GPetites GTPasesFamille dynamineCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculaires