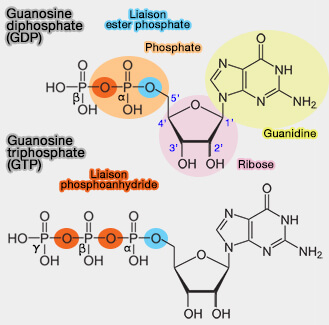
Le domaine G est responsable de changements de conformation dépendant des nucléotides : il agit comme un commutateur moléculaire (switch) entre l'état désactivé lié au GDP (" off ") et l'état activé lié au GTP (" on "), par la réaction :
Les protéines G (ou GNBP, guanine nucléotide-binding protein) sont impliquées dans la transmission de signaux intracellulaires ou entre le milieu extra et intracellulaire.
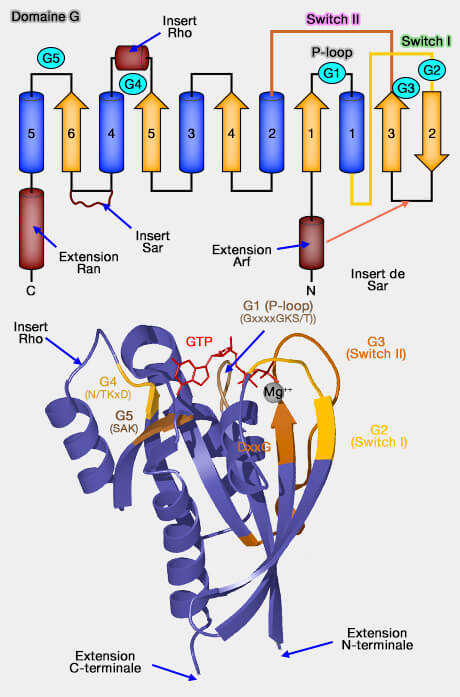
2. Ce domaine G (165 acides aminés) se compose de 5 domaines ou boxes qui sont impliqués dans la liaison avec la guanine et l'ion Mg++.
Les cinq boucles du domaine G sont impliquées dans la liaison du GTP, contrairement à d'autres protéines où les sites de liaison du ligand sont le plus souvent constitués en hélice alpha (An overview of the basic helix-loop-helix proteins 2004) ou de feuillets bêta.
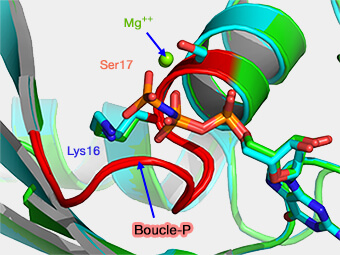
Cette boucle est riche en résidus de glycine et est localisée entre le premier brin du feuillet β (β sheet) et la première hélice α.
Les atomes d'azote de la chaîne principale interagissent étroitement avec les phosphates chargés négativement.
Boucle P (P-loop) de G1
La lysine (K), cruciale pour la liaison au nucléotide, interagit directement avec les oxygènes des phosphate β du GDP (et GTP) et du phosphate γ terminal du GTP.
Le groupe hydroxyle (-OH) de la sérine (S) (S) ou la thréonine (T) contacte l'oxygène du phosphate β et l'ion Mg++.
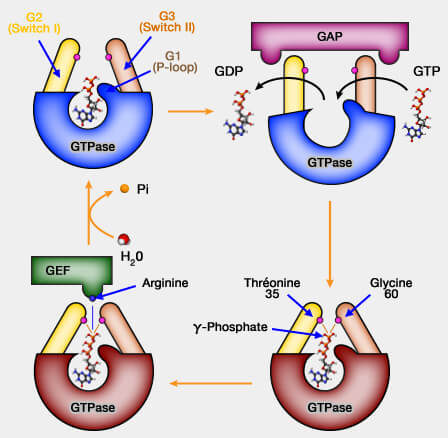
G2
1. G2 comprend le Switch I (32-38), un des interrupteurs, région qui change de conformation lors de l'échange GDP/GTP. On parte aussi de domaine effecteur ou core effector domain : 32-40).
G2 contient la Thr35 (dans H-Ras) qui se lie au γ phosphate terminal du GTP et au Mg++.
Cette thréonine (T) jouerait un rôle dans l’orientation de la boucle qui porte le motif B de Walker (G3) et interviendrait dans la coordination de l’ion magnésium catalytique, mais également dans l’interaction avec les effecteurs.
1. G3 (Switch II) contient le motif DxxG (57-60), qui correspond au motif B de Walker.
Asp57 (acide aspartique) se lierait à la guanine, mais pas forcément au magnésium catalytique, bien qu'il soit très proche.
Glu60 (acide glulutamique dans Switch II, résidues 59–67 chez H-Ras) activerait l'hydrolyse de GTP en GDP, via une liaison hydrogène avec l'oxygène du γ phosphate.
GTPase et changements conformationnels
(Figure : vetopsy.fr)
2. G3 est souvent impliqué dans les interactions effectrices en jouant un rôle dans l'échange des nucléotides par les GEF et la stimulation de l'hydrolyse du GTP par les GAP.
G4
1. G4 contient le motif N/TKxD (116-119)
Ce motif est responsable de la spécificité des GTPases pour les nucléotides à base guanine.
Ce motif apparaît dans une boucle à la fin d’un brin β hydrophobe (souvent le 6ème brin) et aussi impliqué dans la stabilisation de la base du nucléotide.
Les protons du groupement amide de l’asparagine (N) contacte l'oxygène de la purine.
L'asparagine (N) et la lysine (K) conservée stabilisent le site de fixation du nucléotide en interagissant avec les acides aminés de la région G1.
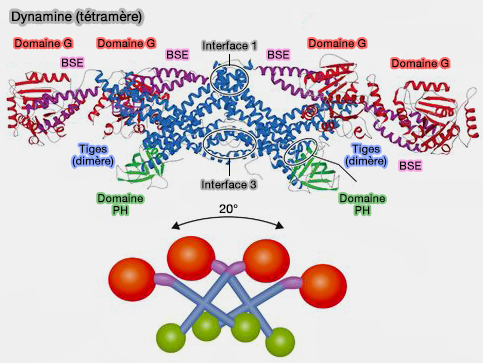
2. G4 et une boucle de stabilisation intervient dans la dimérisation des domaines G, comme dans la dynamine.
G5
1. G5 (guanine cap ou coiffe de guanine), qui n’est pas présent chez toutes les GTPases, mais qui apparaît par exemple dans la dynamine ou Drp1, contient le motif SAX ou SAK - S/CAK/L/T - (145-147), dans une boucle autour du 7ème brin β se liant à la guanine.
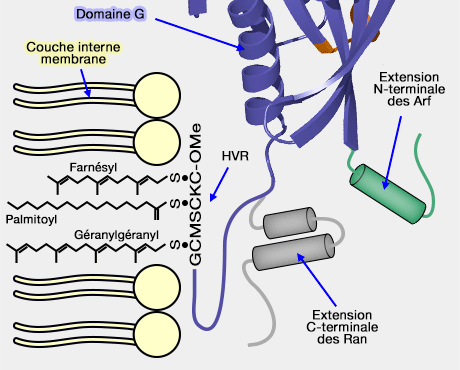
Domaine G, HVR et extensions
(Figure : vetopsy.fr)
L'amine de l'acide aminé central interagit avec l'oxygène de la base de la guanine.
La chaîne latérale de la sérine stabilise l’aspartate et l’asparagine du motif G4 par une liaison hydrogène.
2. L'insert de la famille Rho entre le brin β5 et l'hélice α4, absent dans les membres de cette famille, forme une courte hélice qui sort du reste du domaine G et ne change pas sa position lors de l'échange de nucléotides.
Remarque : attachée à G5, une région hypervariable (HVR) ne faisant pas partie du domaine G (166-188/189 chez Ras) contient un linker et une région C-terminale qui permet d'achever l'attachement membranaire par une modification post-traductionnelle, via des queues lipidiques.
Dimérisation du domaine G
1. L'homodimérisation du domaine G par la poche de liaison aux nucléotides est lié à une hydrolyse coopérative du GTP.
Tétramère de dynamine
(Figure : vetopsy.fr d'après Antonny et coll)
2. Les mécanismes catalytiques peuvent varier suivant les GTPases par rapport à celui de la famille des Ras.
Par exemple, dans la majorité des DSP (Dynamin Superfamily Protein) comme la dynamine ou Drp1, un cation monovalent externe (Na+ ou K+), stabilisé par une sérine conservée (S41) dans la boucle P, est repositionné lors de la dimérisation du domaine G pour annuler la charge négative lors de l'hydrolyse du GTP.






 propriétés de la dynamine).
propriétés de la dynamine).