3. Les transporteurs sont sujets aux modifications post-traductionnelles qui sont essentielles dans la stabilisation, le trafic et l’activité de ces transporteurs :
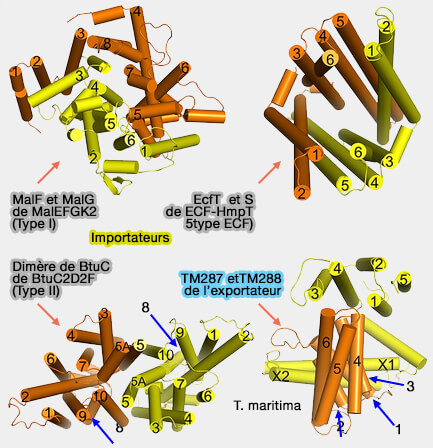
Remarque : chez les procaryotes, les transporteurs sont souvent assemblés à partir de sous-unités protéiques distinctes (2 TMD et 2 NBD) et peuvent être homodimériques ou hétérodimériques.
Parfois, deux ou même trois sous-unités sont fusionnées en une protéine multi-domaine.
4. Les importateurs ABC de type I et II sont dépendants de domaines de liaison au substrat solubles (SBD) ou de protéines de liaison au substrat (SBP) supplémentaires qui capturent le substrat transporté du côté trans et le transmettent aux TMD (liaisons au substrat).
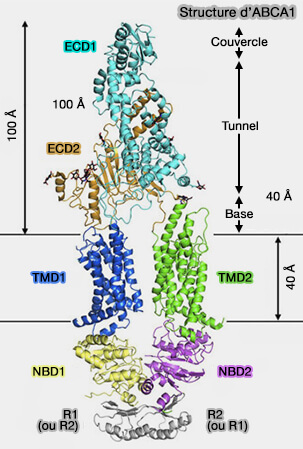
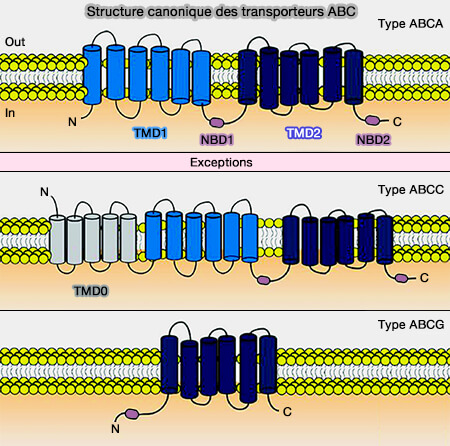
Domaines transmembranaires (TMD)
La structure des TMD (TransMembrane Domain) hydrophobes diffère sensiblement d'une molécule à l'autre.
La voie de translocation est située à l'interface entre les deux TMD (cf. tableau).
Le pore est formé de 10 à 16 hélices α transmembranaires selon le transporteur.
1. Les exportateurs contiennent :
12 TMD pour les transporteurs entiers, quelquefois 17 pour ceux contenant un TMD0,
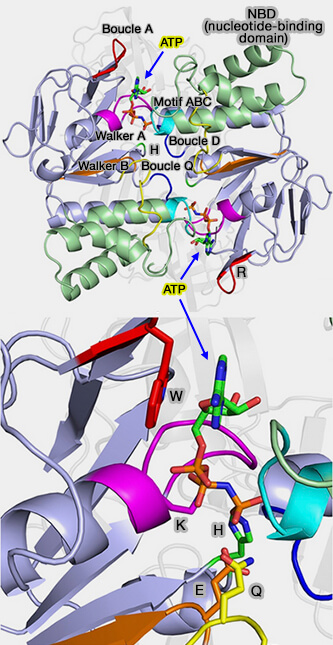
Ce résidu entre en interaction avec l'aspartate (D) conservé de la boucle D, le résidu glutamate (E) du motif Walker B et avec le y-phosphate de l'ATP.
6. La boucle Q, de huit résidus environ, possède, comme son nom l'indique, un résidu glutamine (Q) conservé à son extrémité N-terminale.
Ce résidu est situé à l'interface entre les sous-domaines RecA-like et α-hélicoïdal.
Les changements conformationnels de la boucle Q permettent au résidu de glutamine d'entrer et de sortir du site actif pendant le cycle d'hydrolyse.
Il
permet la formation du site actif lorsque le Mg-ATP est lié et le perturbe une fois que l'ATP est hydrolysé.
La boucle Q est également un site majeur d'interaction avec les TMD.
Orientation des NBD suivant le cycle catalytique
(Figure : vetopsy.fr d'après Beek et Wilkens)
7. La signature ABC (LSGGQ) est caractéristique des protéines ABC, absente des autres ATPases.
Elle est située à l'extrémité N-terminale d'une longue hélice qui dirige la charge positive du dipôle hélicoïdal vers le y-phosphate de l'ATP.
Orientations des NBD
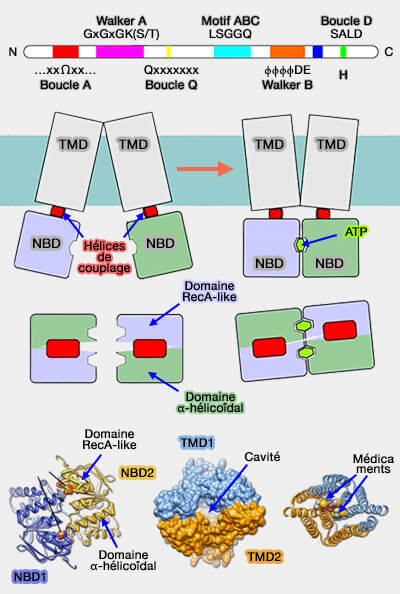
Les deux NBD des transporteurs ABC peuvent adopter des orientations différentes l'une par rapport à l'autre.
1. La conformation fermée plaque les deux NBD l'un contre l'autre grâce aux deux ATP qui, chacune, se lient à un NBD.
La signature ABC d'un monomère est située à proximité des motifs Walker A (boucle P) et B et des boucles A, H et Q de l'autre domaine.
Les sous-domaines RecA et hélicoïdal α de chaque NBD sont tournés l'un vers l'autre.
C'est dans cette conformation que l'hydrolyse peut avoir lieu.
2. La conformation ouverte correspond à une dissociation partielle des deux NBD.
L'hydrolyse de l'ATP en Pi et ADP déstabilise le dimère et permet aux NBD de s'écarter.
Les sous-domaines RecA et hélicoïdal α de chaque NBD s'éloignent l'un de l'autre.
L'énergie chimique de l'hydrolyse de l'ATP est transformée en énergie conformationnelle qui est transmise aux TMD pour favoriser l'accès alterné de la voie de translocation du substrat des deux côtés de la membrane.
a. L'hélice de couplage est une hélice α courte située dans une protubérance cytoplasmique du TMD qui s'encoche dans une rainure d'un monomère NBD.
La région des NBD qui interagit avec l'hélice de couplage des TMD contient la boucle Q.
Cette rainure est située exactement à l'interface entre le sous-domaine hélicoïdal α et le sous-domaine RecA, qui tournent l'un vers l'autre en réponse à la liaison de l'ATP pour l'hydrolyse de l'ATP.
b. De cette manière, chaque NBD est connecté à un TMD, et les changements de conformation dans les NBD peuvent être transformés en changements conformationnels dans les TMD.
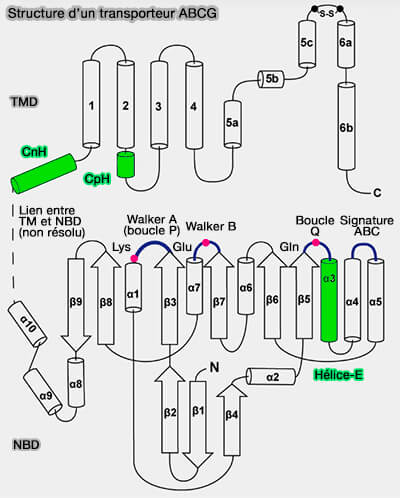
2. Dans certains transporteurs, on trouve deux autres hélices comme dans les transporteurs ABCG :





 liaisons au substrat).
liaisons au substrat).