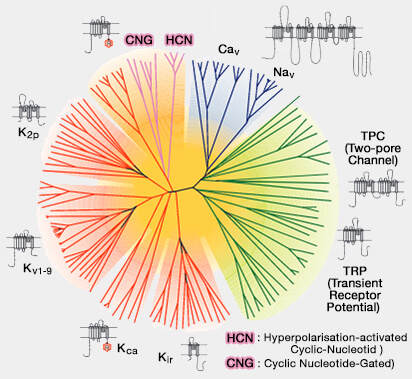
Toutefois, malgré la présence du domaine de capteur de tension commun à tous les membres de cette famille, ils n'y sont pas sensibles, mais sont déclenchés par la concentration intracellulaire de l'AMPc ou du GMPc.
La superfamille des VGL
(Figure : vetopsy.fr d'après Yu et Catterall)
2. Ces canaux sont constitués de quatre sous-unités, correspondant parfois à 4 domaines d'une même protéine, comme pour le canal sodium voltage-dépendent (NaV).
3. Les canaux CNG sont des canaux cationiques non sélectifs qui se trouvent dans les membranes de divers tissus et de cellules et jouent un rôle important dans
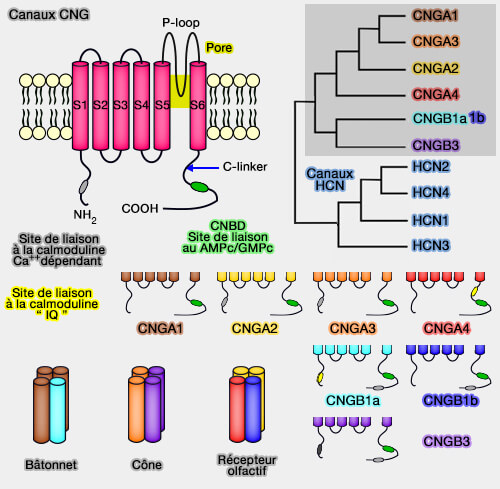
Les canaux CNG, formés de 4 sous-unités, ont d'abord été trouvés dans les photorécepteurs, puis dans les cellules olfactives, et enfin dans d'autres cellules nerveuses ou non.
Chez les mammifères, une famille de six gènes codent pour :
Bien que possédant un capteur de tension, les canaux CNG ne sont pas bloqués par la tension, une propriété essentielle pour une phototransduction et une transduction olfactive appropriées.
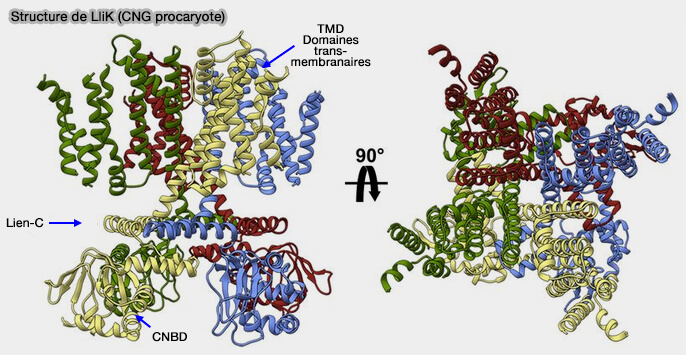
Structure de LliK (CNG procaryote)
(Figure : vetopsy.fr d'après James and coll)
Liaison VSD-pore non " permutée "
(Figure : vetopsy.fr d'après James and coll)
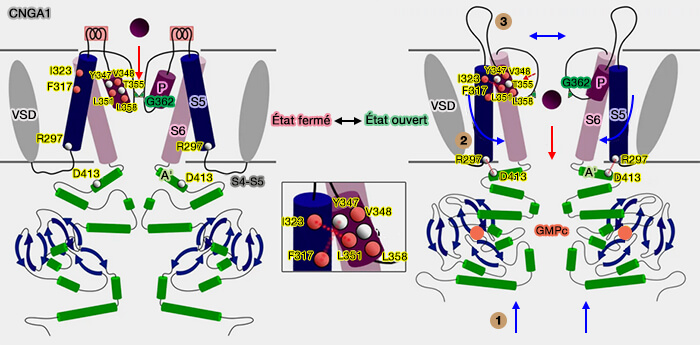
Cette boucle P permet la sélectivité ionique du canal CNG et détermine le diamètre du pore central pour laisser passer les ions en fonction de leur diamètre.
Fermée, cette porte empêche les ions de pénétrer dans le canal.
Des changements conformationnels des quatre sous-unités permet l'ouverture du canal.
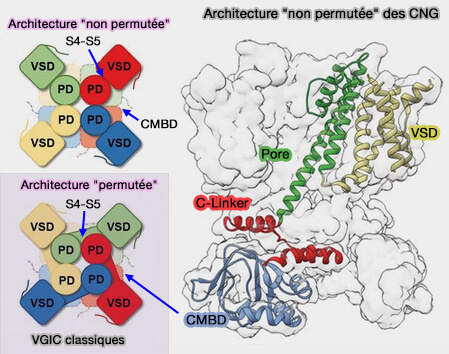
3. La liaison entre le VSD et le domaine du pore est particulière.
Dans les canaux CNG, le VSD interagit uniquement avec le domaine du pore de la même sous-unité dans une configuration dite " non permutée " par des interactions hydrophobes et des liaisons hydrogène entre les extrémités extracellulaires de S1 et S5 et l'hélice des pores, et entre les extrémités intracellulaires de S4 et S5.
Extrémité C-terminale
Les deux extrémités C- et N-terminales sont intracellulaires et contiennent des régions fonctionnelles pour la régulation du canal.
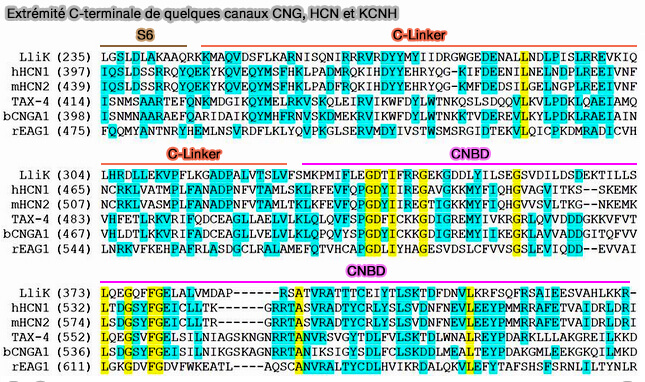
Extrémité C-terminale de quelques canaux CNG, HCN et KCNH
(Figure : vetopsy.fr d'après James and coll)
L'extrémité C-terminale comporte deux régions essentielles au fonctionnement du canal.
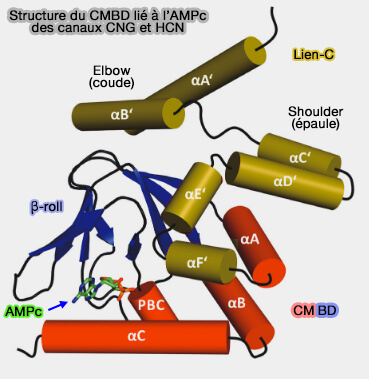
1. Le site de liaison du nucléotide cyclique ( domaine CNBD : Cyclic nucleotide-binding domain), i.e. l'AMPc pour les récepteurs olfactifs, GMPc pour les photorécepteurs, est classique et retrouvé dans d'autres protéines.
Le rouleau β est entouré par l'hélice α N-terminale, une courte hélice α interne située entre les brins β6 et β7, désignée comme la cassette de liaison au phosphate (phosphate-binding-cassette : PBC) et deux hélices α (αB et αC) situé à l'extrémité C-terminale.
Le nucléotide cyclique se lie à l'intérieur d'une cavité formée par certaines parties du rouleau β, l'hélice PBC et l'hélice αC.
2. Le lien-C (C-linker) est une région d'environ 80 acides aminés qui relie le CNBD au segment S6, et est essentiel au déclenchement du canal.
L'hélice A' est connectée à S6 via un simple lien entre 2 acides aminés. Cette connexion directe est cruciale pour l'activation.
Les hélices A'B' s'étendent à un angle de 25 ° par rapport au plan de la membrane et sont prises en sandwich entre le domaine transmembranaire (TMD) et le reste de l'extrémité C-terminale.
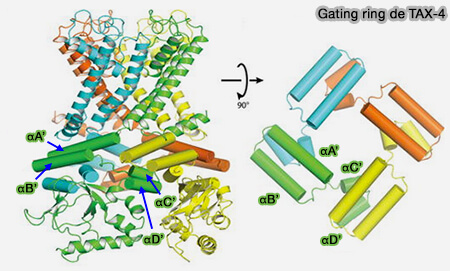
Gating ring de TAX-4 (Figure : Li and coll)
Les deux hélices A'B' forme un motif hélice-coude-hélice antiparallèle qui entre en contact avec les hélices αC′ and αD′ de la sous-unité voisine, appelé elbow-shoulder (coude-épaule) ou " gating ring ".
D'autres interactions entrent en jeu comme des liaisons hydrogène et un pont salin entre les résidus K472 dans l'hélice αB′ et E502 dans αD′ de la sous-unité voisine.
La tétramérisation du canal est due à deux interfaces: la première dans Des interactions hydrophobes sont aussi présentes entre le domaine du pore d'une sous-unité avec S6 de la sous-unité voisine(Structure of a eukaryotic cyclic nucleotide-gated channel 2017).
Les hélices A'B' interagissent avec le domaine transmembranaire et maintiennent S6 dans un état ouvert. Grâce à ces interactions, les hélices A'B' transmettent directement les changements de conformation du CNBD au TMD et finalement au filtre de sélectivité.




 transduction visuelle) et l'olfaction (
transduction visuelle) et l'olfaction (