Traitement rétinien de la vision
Photorécepteurs : phototransduction
Citation
« Chacun a ses lunettes ; mais personne ne sait
au juste de quelle couleur en sont les verres. »
Alfred de Musset
Documentation web
- La vision (
 infos)
infos) - La vision dans le monde animal et chez l'homme
( infos)
- Tout savoir sur les couleurs
( infos)
- L'oeil humain avec logiciel
( infos)
- Voies visuelles ( infos) &
( infos) & ( infos)
- Les illusions optiques
( infos)
- Photothèque d'ophtalmologie
( infos)
- Vision
- Neurophysiologie
de la vision
- La lumière et ses propriétés
- Formation de l'image dans l'oeil
- Traitement rétinien de l'image
- Voies optiques
- Communication visuelle
- Communication chez l'homme
![]()
Les photorécepteurs
sont les cellules qui transforment l'énergie lumineuse, c'est-à-dire
électromagnétique, en énergie électrique : c'est
la transduction visuelle (![]() infos).
infos).
Les photorécepteurs, contenus dans la couche neuro-épithéliale rétinienne, sont de deux types et correspondent à deux systèmes différents :
- les bâtonnets (un seul type) ;
Ces photorécepteurs font partie du système scotopique (en grec, skotos, obscurité) : ils travaillent à de faibles luminosités (vision crépusculaire), ne sont pas sensibles à la couleur et détectent grossièrement les formes.
- les cônes qui sont représentés par 3, et
maintenant 4 types différents selon le pigment qu'ils contiennent
(
infos).
Ces photorécepteurs font partie du système photopique (en grec, photos, lumière) : il permettent la vision des couleurs et détectent également les formes.
Les réactions, au niveau des photorécepteurs, sont de deux ordres se produit en deux étapes :
- la première est photochimique,
- la deuxième est électrophysiologique.
Etape biochimique
Rétinal (
infos)
![]()
 C'est grâce au rétinal, molécule photosensible (aldéhyde
de la vitamine
A), que les photorécepteurs transforment la lumière en
signaux électriques.
C'est grâce au rétinal, molécule photosensible (aldéhyde
de la vitamine
A), que les photorécepteurs transforment la lumière en
signaux électriques.
Le rétinal se combine à des protéines, nommées opsines, pour former quatre types de pigments différents qui absorbent différentes longueurs d'ondes de la lumière visible.
Ce sont les cellules de la couche pigmentée qui capte et stocke la vitamine A, la transforme et la transfère aux photorécepteurs.
Le rétinal peut adopter diverses structures tridimensionnelles appelées isomères.
- Lié à une opsine, il est sous forme rétinal 11-cis (entortillé).
- Quand il absorbe des photons, c'est-à-dire qu'il est frappé par la lumière, il passe en forme trans (redressée) et il se détache de l'opsine.
![]()
C'est là le seul stade qui dépend de la lumière
! Ce phénomène déclenche tout le processus
de la phototransduction.
Rhodopsine (
infos)
La phototransduction la mieux connue est celle des bâtonnets qui contiennent la rhodopsine au niveau des disques empilés du segment externe.
") La
rhodopsine (du grec rhodos, rose, et opsis, vision) est un pigment de couleur
pourpre, disposée au travers des membranes des disques des segments
externes. Elle est constituée par :
La
rhodopsine (du grec rhodos, rose, et opsis, vision) est un pigment de couleur
pourpre, disposée au travers des membranes des disques des segments
externes. Elle est constituée par :
- une partie protéique (l'opsine qui est la scotopsine dans les bâtonnets),
- le 11-cis-rétinal (cf. plus haut).
La rhodopsine est composé de 348 acides aminés et son gène codant est situé sur le chromosome 3.
Bien que la rhodopsine absorbe la lumière de tout le spectre visible, elle absorbe surtout les longueurs d'onde de 497 nm (lumière verte).
Lorsque la rhodopsine est frappée par la lumière, les deux composants se séparent : c'est ce que l'on nomme la décoloration de la rhodopsine.
Le tout-trans-rétinal est absorbé par l'étroit espace sous-rétinien compris entre l'épithélium pigmentaire et les photorécepteurs. Cet espace est rempli par un liquide gélatineux qui contient la protéine de liaison intra-rétinienne (IRPB : intra-retinoïd binding protein) qui conduit le rétinal jusqu'à l'épithélium pigmentaire où il est retransformé en 11-cis pour retourner dans les segments externes des photorécepteurs. Il se lie à l'opsine pour reconstituer la rhodopsine .
![]()
La carence en vitamine
A perturbe, tout d'abord, la vision
nocturne pour entraîner une cécité complète
par du segment externe du photorécepteur.
La rhodopsine est activée par la lumière (1).
")
La rhodopsine activée (2) dévoile un site qui fixe une protéine G, la transducine composée de trois sous-unités (a, b et g). La transducine activée, en fixant un GTP, libère sa fraction a. (3) .
La fraction a de la transducine active à son tour la phosphodiestérase, composée elle-même de trois sous-unités (4), qui hydrolyse le GMP cyclique (Guanosine 3’-5’ monophosphate cyclique) en 5'-GMP (5).
Ce GMPc, fixé sur les canaux Na+ du segment externe du photorécepteur, est responsable de sa béance (A). Quand il est hydrolysé, le canal se ferme (B), provoquant une hyperpolarisation membranaire, qui diminue la quantité de neuromédiateur libéré dans les terminaisons synaptiques au contact avec les cellules bipolaires.
Étape électrophysiologique
Excitation des bâtonnets
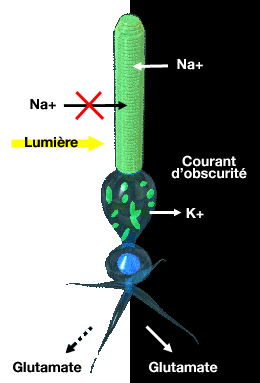 A
l'obscurité, les canaux Na+ du segment externe du photorécepteur
sont ouverts et laisse passer librement les ions, grâce à une
forte concentration de GMPc.
A
l'obscurité, les canaux Na+ du segment externe du photorécepteur
sont ouverts et laisse passer librement les ions, grâce à une
forte concentration de GMPc.
Les ions K+ sortent du segment interne vers l'espace extracellulaire. Ce déplacement provoque un courant appelé" courant d'obscurité "
Ce courant entrant dépolarise le photorécepteur avec un potentiel de membrane intermédiaire autour de -40 mV.
Les bâtonnets peuvent ainsi libérer plus ou moins continuellement leur neurotransmetteur au niveau de leurs synapses avec les cellules bipolaires.
Les bâtonnets, contrairement aux différents cônes, ne sont sensibles qu'à une seule longueur d'onde (aux environs de 500 nm, c'est-à-dire aux couleurs bleu-vert).
C'est pour cette raison que les affichages des appareils électriques qui doivent être vus dans l'obscurité (comme le cadran du réveil par exemple) sont dans les tons verts.
Certains préfèreraient des tons rouges vifs pour stimuler les cônes et laisser les bâtonnets au repos pour une meilleure vision nocturne.
La lumière ferme les canaux Na+ ce qui a pour effet d'hyperpolariser les photorécepteurs (cf. plus haut) : le potentiel de membrane tend vers le potentiel d’équilibre du K+ à -70 mV, ce qui inhibe la libération de neurotransmetteur.
![]()
En règle générale, dans les systèmes synaptiques,
c'est l'inverse qui se produit !
![]()
Le système visuel réagit à des changements d'éclairement
: une hyperpolarisation comme une dépolarisation permet de les détecter.
Les cônes, les bâtonnets, et les cellules horizontales agissent les unes sur les autres par des potentiels électriques locaux gradués ou potentiels récepteurs par l'intermédiaire de la libération également graduée du neurotransmetteur.
Par contre, les cellules amacrines et les cellules ganglionnaires produisent des potentiels d'action par car leurs axones propagent des influx nerveux dans les nerfs optiques (II).
Excitation des cônes
Chez l'homme
Les pigments visuels des cônes, appelés photopsines, se dégradent et se régénèrent de la même manière que la rhodopsine.
Les photopsines sont formés de rétinal et d'opsines différentes de la scotopsine des bâtonnets.
 Par
contre, leur seuil d'excitation est bien plus élevé que celui
des bâtonnets,
ce qui explique que les cônes ne réagissent qu'à la
lumière intense.
Par
contre, leur seuil d'excitation est bien plus élevé que celui
des bâtonnets,
ce qui explique que les cônes ne réagissent qu'à la
lumière intense.
Les cônes contiennent une des trois catégories d'opsines sensibles. Le nom des cônes fait référence aux couleurs, c'est-à-dire les longueurs d'onde, qu'ils absorbent de préférence.
- Les cônes bleus (ou S, supra frequency) réagissent surtout aux longueurs d'onde de 430 nm (5% des cônes) ;
Le pigment des cônes S possède 348 acides aminés comme la rhodopsine dont environ 40% en commun. Son gène codant est situé sur le chromosome 7.
La ressemblance entre le pigment des cônes S et la rhodopsine fait penser que cette dernière dérive du premier.
- les verts (ou M, medial frequency), à des longueurs d'onde de 530 nm ;
Le pigment des cônes M possède 364 acides aminés. Son gène codant est situé sur le chromosome X.
- les " rouges " (ou L, low frequency), en réalité jaunes-vert, aux longueurs d'onde de 580 nm ;
Le pigment des cônes L possède 364 acides aminés et ne diffère que de 15 acides aminés du pigment des cônes M. Son gène codant est situé sur le chromosome X, devant celui du cône L qui sont donc l'un derrière l'autre sur le bras long du chromosome X.
Leur origine est commune et leur divergence à partir d'un pigment commun n'a eu lieu que récemment (30 millions d'années).
- un quatrième type de cône vient
d'être découvert : il est sensible à
la mélanopsine (
infos).
Les longueurs d'ondes des différents cônes
sont les plus à même d'induire le changement de configuration
du rétinal et sa séparation de l'opsine. Autrement dit, chaque
photopsine induit un milieu électrique particulier.
![]()
 L'oeil
de l'homme peut reproduire toutes les couleurs en mélangeant les
pigments bleus, vert et rouge (théorie de trichromie de Young-Helmholtz).
L'oeil
de l'homme peut reproduire toutes les couleurs en mélangeant les
pigments bleus, vert et rouge (théorie de trichromie de Young-Helmholtz).
C'est le physicien Thomas
Young (1773-1829) qui établit cette théorie en 1802 confirmée
par le physiologiste Hermann Ludwig von Helmholtz (1821-1894) qui inventa
entre autres l'ophtalmoscope (![]() infos).
infos).
- C'est ce principe qui est utilisé dans les tubes cathodiques
(
 Wiki).
Wiki).
- Par contre, ce que l'on nomme couleurs primaires sont le bleu, le jaune
et le rouge (
infos).
La perception des couleurs intermédiaires résulte
de l'activation simultanée, mais plus ou moins prononcée d'un
type de cônes (![]() infos).
infos).
La lumière jaune stimule les cônes rouges et les cônes verts, mais si les rouges sont plus stimulés que les verts, nous voyons de l'orange. Si tous les cônes sont stimulés de manière égale, nous voyons du blanc.
La perception de la brillance et de la saturation des couleurs (du rouge par opposition au rose par exemple) est aussi liée au degré de stimulation de chaque type de cônes.
Les possibilités de discrimination des couleurs sont amoindries
chez une faible proportion d'individus que l'on qualifie de daltoniens (![]() infos).
infos).
Chez les animaux (
infos)
") La sensibilité chromatique, capacité à distinguer les
couleurs, dépend des variétés de cônes.
La sensibilité chromatique, capacité à distinguer les
couleurs, dépend des variétés de cônes.
Les études physiologiques sur la sensibilité chromatique montrent que nos carnivores domestiques (chat et chien) sont susceptibles de distinguer plusieurs couleurs.
![]()
Ils sont dichromates, contrairement aux humains qui sont trichromates (![]() infos).
infos).
Leurs performances sont nettement supérieures dans les bleus et les jaunes, tandis que leur rétine semble peu sensible à la lumière rouge qui leur paraît foncé et le vert se confond avec le blanc : une pelouse de printemps sera blanchâtre et un rosier en fleur aura l'aspect d'un buisson blanc parsemé de tâches noirâtres.
![]()
La nuit, les chiens et les chats voient, comme nous, en noir et blanc :
les cônes deviennent insensibles en lumière faible.
Cellules bipolaires et ganglionnaires : champs rÃĐcepteurs
Vision![]() Structure
oculaire
Structure
oculaire![]() Perception visuelle
Perception visuelle![]() Vision
du chien
Vision
du chien![]() Vision
du chat
Vision
du chat
![]() Neurophysiologie de la vision
Neurophysiologie de la vision![]() PropriÃĐtÃĐs de la lumiÃĻre
PropriÃĐtÃĐs de la lumiÃĻre
![]() Formation de l'image dans l'oeil
Formation de l'image dans l'oeil![]() Traitement rÃĐtinien de l'image
Traitement rÃĐtinien de l'image![]() RÃĐtine
RÃĐtine
![]() Photorécepteurs
Photorécepteurs![]() Cellules
bipolaires et ganglionnaires
Cellules
bipolaires et ganglionnaires![]() Cellules
horizontales et amacrines
Cellules
horizontales et amacrines
![]() Voies optiques
Voies optiques![]() Communication
visuelle
Communication
visuelle
![]() Communication
visuelle du chien
Communication
visuelle du chien![]() Communication
visuelle du chat
Communication
visuelle du chat
- Lignereux Y - Eléments d'anatomie ophtalmologique du chien - Pratique médicale et chirurgicale de l'animal de compagnie , 32 (supplément), 1997
- Kahle W. - Anatomie, Tome 3 Système nerveux et organes des sens - Flammarion Médecine-Sciences, 423 p., 2007
- Marieb E. N. - Anatomie et physiologie humaines - De Boeck Université, Saint-Laurent, 1054 p., 1993
- Rosenzweig M.R., Leiman A.L., Breedlove S.M. - Psychobiologie - DeBoeck Université, Bruxelles, 849 p., 1998
- Purves D., Augustine G.J., Fitzpatrick D., Katz L.C., Lamantia A-S, McNamara J.O., Williams S.M. - Neurosciences - De Boeck, 800 p., 2003
- Kolb B., Whishaw I. - Cerveau et comportement - De Boeck, 646 p., 2002
- Bear M.F., Connors B.W., Paradiso M.A. - Neurosciences : à la découverte du cerveau - Masson-Williams, 654 p, 1997
- Gazzaniga, Ivry, Mangun - Neurosciences cognitives - De Boeck, 585 p., 2001
- Rodieck R. W. - La vision - De Boeck, 562 p., 2003
- Habib M. - Bases neurobiologiques des comportements - Masson, 276 p., 1998
- Clerc B. - Ophtalmologie vétérinaire - Editions du Point vétérinaire, Maisons-Alfort, 664 p;, 1997
- Clerc B. - L'oeil du chat : particularités anatomiques et physiologiques - Pratique médicale et chirurgicale de l'animal de compagnie , 27 (supplément), 1992
- Simon M. - Le fond d'oeil normal chez les carnivores domestiques - Receuil de médecine vétérinaire, 165 (3), 1989
- Chaudieu G. - Eléments d'études simples des affections du fond d'oeil chez les carnivores domestiques - Pratique médicale et chirurgicale de l'animal de compagnie , 31, 1996
- Sauret (J) et Machie (A) - L'oeil des carnivores et ses annexes - Rec Med Vet, 165 (3), 175-186, 1989