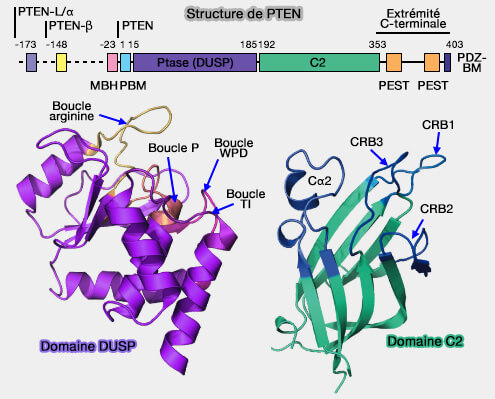
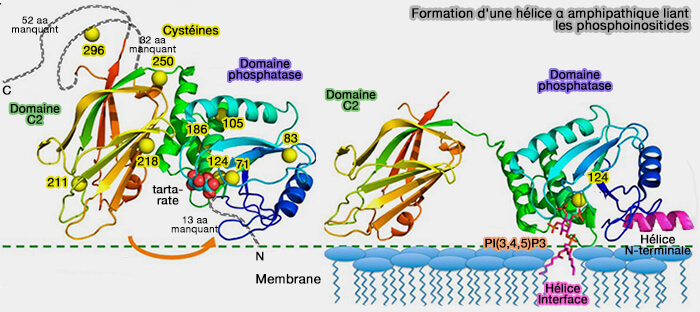
1. L'extrémité N-terminale comprend un motif PBM (Phosphoinositide Binding Motif), i.e. domaine de liaison à PI(4,5)P2 ou PIP2, (1-MTAIIKEIVSRNKRR-15) qui détermine la up-régulation de l'activité de PTEN par un mécanisme allostérique.
Des mutations des résidus de cette poche (S10N, G20E, L42R et F90S) diminue l'affinité pour de PI(4,5)P2 et provoque l'apparition de tumeur en diminuant l'affinité membranaire de PTEN.
Formation d'une hélice α amphipathique liant les phosphoinositides
(Figure : vetopsy.fr d'après Wei et coll)
2. L'attachement à la membrane s'effectue également via le domaine PDZ C-terminal qui se lie à ses homologues des protéines membranaires.
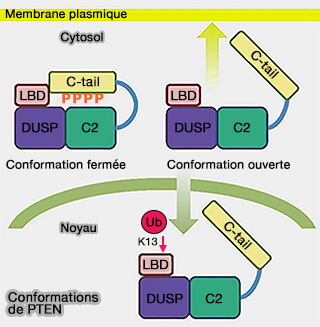
que l'extrémité C-terminale ne soit pas phosphorylée,
que K13 ne soit pas ubiquitinée, l'ubiquitination la transportant vers le noyau ( régulations de PTEN).
Domaine phosphatase
Le domaine lipidique phosphatase, résidus 15-185, lié à la protéine tyrosine phosphatase (PTP), est constitué d'un feuillet β à cinq brins β pris en sandwich entre 6 hélices α, i.e. 2 d'un côté, 4 de l'autre.
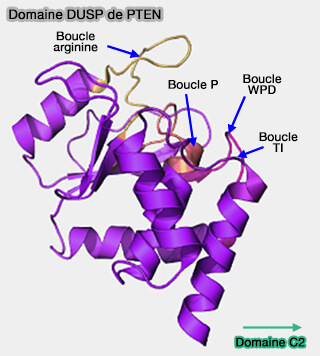
Structure du domaine
Domaine DUSP de PTEN
(Figure : vetopsy.fr d'après Masson et coll)
Le noyau du domaine phosphatase, qui contient quatre brins β parallèles et une hélice α avec la boucle P catalytique reliant l'un des brins β et l'hélice α, est conservé.
1. Le site actif de PTEN comprend en plus - trois boucles, qui forment une poche plus profonde et plus large que celle des protéines tyrosine phosphatases courantes, i.e. ce qui permet au groupe de tête volumineux de PI(3,4,5)P3 ou PIP3 d'accéder au centre actif de l'enzyme, i.e. C124.
la boucle WPD (résidus 88-98), dans lequel le résidu Asp fonctionne à la fois comme un acide et une base lors de la catalyse,
la boucle TI (résidus 160-171) qui contient une paire de thréonine et d'isoleucine, qui élargit la poche,
la boucle P.
2. Cette poche catalytique est fortement chargée positivement,plus que les autres PI-phosphatases, comme les myotubularines (MTM), ce qui est en accord avec sa préférence pour PI(3,4,5)P3, le PI chargé le plus négativement.
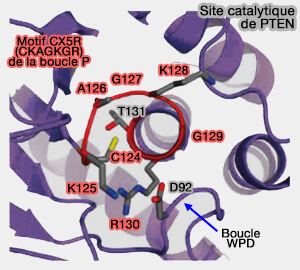
Site catalytique de PTEN
(Figure : vetopsy.fr d'après Manford et coll)
La boucle P catalytique contient le motif CX5R, dont la séquence centrale est ici, CKAGKGR forme la poche ou la fente catalytique de PTEN, qui est adapté à la préférence de PTEN pour PI(3,4,5)P3.
Les sept groupes amide consécutifs du motif CX5R sont tous tournés vers l'intérieur, conjointement avec le groupement guanidinium de l'arginine (R).
Cette conformation permet de placer correctement le groupement phosphate du substrat pour l'attaque nucléophile par l'atome Sγ de la cystéine catalytique.
Par contre, le domaine catalytique de SAC1, bien qu'il contienne le motif CX5R, est différent ( structure de SAC1).
2. Le mécanisme est le suivant.
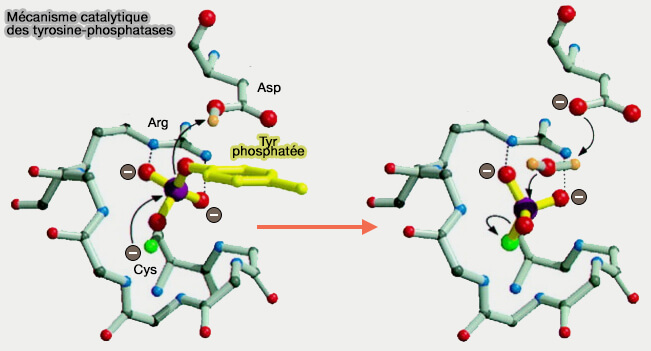
a. L'anion thiolate de la cystéine du site actif (C124 de PTEN) effectue une attaque nucléophile sur le phosphate du substrat, sur la figure, un phosphoryl-tyrosyle.
Cela provoque la formation d'un intermédiaire phospho-enzyme, i.e. un cystéinyle phosphoenzyme covalent intermédiaire.
L'acide aspartique, (D92 de la boucle WPD de PTEN), agissant comme un acide général, donne un proton au groupe phosphate, éjectant la chaîne latérale du substrat. Les liaisons hydrogène entre les atomes d'oxygène du phosphate et les atomes d'azote du groupe favorisent la liaison du phosphate et stabilisent l'état de transition.
b. Agissant comme base générale, l'aspartate active une molécule d'eau pour une attaque nucléophile sur l'atome de phosphore. La liaison phosphore-soufre est hydrolysée, régénérant ainsi l'enzyme native.
Mécanisme catalytique des tyrosine-phosphatases
(Figure : vetopsy.fr d'après Denu et coll)
La mutation de G129, trouvée dans le syndrome de Cowden, inhibe l'activité phosphatase sur les lipides, mais pas sur les protéines,
Les mutations de Y138 ont le rôle inverse, i.e. inhibent l'activité phosphatase sur les protéines mais pas sur les lipides, ce qui a permis de déterminer leurs contributions respectives sur les fonctions de PTEN.
Domaine C2
Le domaine C2 de type II (résidus 192-353), indépendant du calcium, est un domaine structural protéique constitué d'un noyau β-sandwich composé de deux feuillets β antiparallèles et de deux courtes hélices α près de l'interface.
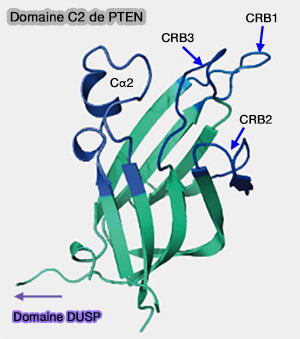
Domaine C2 de PTEN
(Figure : vetopsy.fr d'après Masson et coll)
La boucle Cα2 est plus longue dans le domaine de PTEN que dans des structures homologues, et cette boucle joue un rôle :
dans les interactions entre les deux domaines PTEN.
Le domaine C2 est impliqué dans le ciblage des protéines et la fixation non-spécifique des lipides dans les membranes cellulaires, en particulier par des boucles de liaison au Ca++ (CBR1/2) et une boucle de liaison à la membrane (CBR3).
Dans PTEN, ces boucles diffèrent des boucles Ca++-dépendantes en ayant perdu les résidus responsables ce cette liaison.
Par contre, la boucle CRB3 a une charge positive de +5 et l'interaction à la membrane se fait à travers elle, bien que d'autres boucles puissent être invoquées.
1. Les domaines phosphatase et C2 sont étroitement serrés l'un contre l'autre, partageant une interface étendue (1400 Å2) enrichie de résidus hydrophobes et aromatiques, et un réseau de liaisons hydrogène interdomaine.
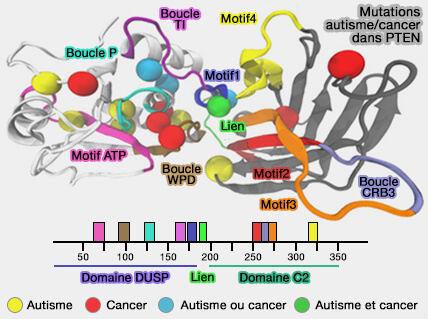
Mutations autisme/cancer dans PTEN
(Figure : vetopsy.fr d'après Smith et coll)
1. L'extrémité C-terminale contient de nombreux sites de phosphorylation essentiels pour régler l'activité et les fonctions de PTEN ( régulations de PTEN).
Conformations de PTEN
(Figure : vetopsy.fr d'après Nguyen et coll)
a. Par exemple, les deux séquences PEST, régions riches en proline (P), en acide glutamique (E), en sérine (S) et en thréonine (P), sont des sites de phosphorylation, en particulier les sérines 380, 382, 383 et 386.




 mutations de PTEN).
mutations de PTEN).