


Fusion membranaire : mécanisme
Formation du pore de fusion (3)
Rôles des protéines associées aux SNARE
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Transport membranaire
- Transports membranaires sans mouvements membranaires
- Transports membranaires avec mouvements membranaires : trafic vésiculaire
- Protéines membranaires
- Moteurs moléculaires
- Voies de signalisation

Le mécanisme de la formation du pore de fusion est dépendant :
Les protéines associées au complexe SNARE sont :
- les Protéines d'attache (tethering protein) comme le complexe HOPS,
- les protéines Sec1/Munc18-like (SM) et Munc13,
- les synaptotagmines par leurs domaines C2 dans les vésicules synaptiques (
 mécanisme général des synaptotagmines).
mécanisme général des synaptotagmines).

Toutes ces protéines sont étudiées dans des chapitres spéciaux.
Protéines associées aux SNARE et formation du pore de fusion
1. Le site de fusion entre deux vacuoles de levure est situé près du bord de la zone de contact, i.e. placé asymétriquement ( pore de vertex).
L'addition du complexe HOPS conduit à l'ouverture des pores (A tethering complex drives the terminal stage of SNARE-dependent membrane fusion 2017).
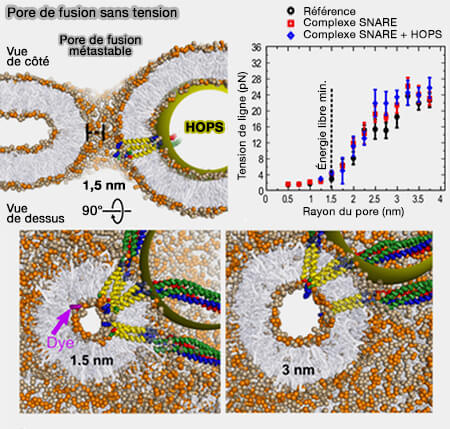
(Figure : vetopsy.fr d'après D'agostino et coll)
1. La sphère jaune imite une perturbation stérique sur le site de fusion imposée par la liaison HOPS à l'un des complexes SNARE.
- La partie supérieure de la figure représente une coupe transversale de ce pore métastable de 1,5 nm de diamètre en vue latérale.
- Ces pores restent largement sphériques malgré l'hétérogénéité du site de fusion.
La vue de dessus montre également une molécule de colorant fluorescent en violet (dichlorofluorescéine) pour illustrer son échelle par rapport au pore de 1,5nm.
2. L'ouverture des pores de fusion peut être stimulée de la même manière lorsque HOPS est remplacé par d'autres protéines artificielles de liaison à SNARE, i.e. les seules propriétés spécifiques requises pour la stimulation de la fusion sont que :
- ces ligands artificiels ajoutent un volume similaire au complexe SNARE,
- ils se lient à proximité du site de fusion membranaire, sur ou proche du domaine SNARE.
La déformation du diaphragme d'hémifusion abaisserait la barrière énergétique pour l'ouverture des pores, de 67 à 34 kBT, et fournirait la force motrice.
3. Les protéines de liaison, i.e. protéines Sec1/Munc18-like (SM) et Protéines d'attache (tethering protein), i.e. volumineuses protéines, sont essentielles pour surmonter la barrière énergétique finale à la formation des pores de fusion par plusieurs mécanismes.
a. Elles conduisent les SNARE et le site de fusion vers le bord de la zone de contact, et de ce fait concentrent les SNARE à ce niveau.
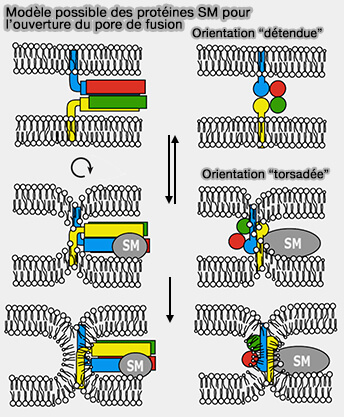
(Figure : vetopsy.fr d'après Pieren et coll)
b. Le volume accru des protéines non-SNARE au site de fusion augmente la courbure membranaire imposée à la tige et anticipe partiellement celle d'un pore de fusion ultérieur (Teardrop Shapes Minimize Bending Energy of Fusion Pores Connecting Planar Bilayers 2015).
c. Par leur volume, elles éloigneraient les complexes SNARE du site de fusion et généreraient une force de traction supplémentaire sur les extrémités C-terminales de la tige indentée.
Remarque : les protéines de liaison pourraient aussi dicter un positionnement torsadé des domaines SNARE au site de fusion (The SM protein Vps33 and the t-SNARE Habc domain promote fusion pore opening 2010).
- De ce fait, ils pourraient permettre au complexe SNARE de se fermer jusqu'à un demi-tour de plus qu'il ne le ferait normalement dans un environnement relaxé (détendu), i.e. lorsque le positionnement en rotation des domaines SNARE n'est pas limité par les protéines associées.
- Ce mécanisme fournirait une force de traction supplémentaire sur les domaines transmembranaires (TMD).
Les SNARE et les protéines chaperons formeraient ensemble la machinerie de fusion.
Synaptotagmine et formation du pore de fusion
Dans un des modèles de fusion, décrit dans le chapitre sur la synaptotagmine, Syt1 interagit au niveau de la surface incurvée négativement qui est présente après la formation d'un intermédiaire d'hémifusion (Conserved arginine residues in synaptotagmin 1 regulate fusion pore expansion through membrane contact 2021).
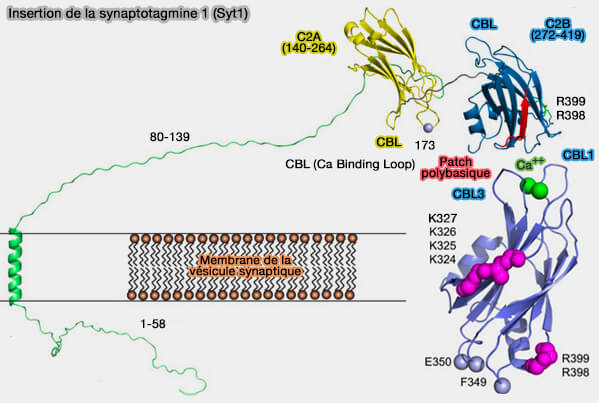
(Figure : vetopsy.fr d'après Nyenhuis et coll)
1. Après l'amarrage à la bicouche et en présence d'ATP/Mg++, le complexe SNARE, i.e. syntaxine 1 (Stx1), SNAP-25, synaptobrévine 2/VAMP2, et le domaine C2B de la synaptotagmine 1 (Syt1) favorisent le mélange partiel des lipides par la fermeture à glissière ou par le contact avec PI(4,5)P2 ou PIP2 de la membrane plasmique au niveau de l'apex d'arginine et de la face polybasique.
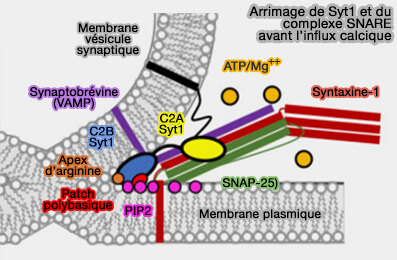
(Figure : vetopsy.fr d'après Nyenhuis et coll)
2. Lors de l'afflux calcique, le domaine C2A de Syt1 s'insère dans la membrane de la vésicule synaptique, la phosphatidylsérine (PS) et C2B se réoriente, liant les boucles de liaison au Ca++ à un troisième point de contact membranaire, i. e. apex d'arginine, patch polybasique et boucles de liaison au Ca++.
a. Le domaine C2B de Syt1 aide à assurer la transition lors d'un événement de fusion vers un pore complètement ouvert :
- soit en abaissant la barrière énergétique pour l'expansion des pores,
- soit en empêchant le système d'être piégé dans un état intermédiaire précoce lors de l'ouverture des pores (Membrane fusion: a structural perspective on the interplay of lipids and proteins 2003).
En affaiblissant ou en éliminant le contact membranaire de l'apex d'arginine, i.e. R398 et R399 avec les mutations RQ ou RQRQ, le domaine C2B n'assume plus l'orientation correcte à travers le côté cytoplasmique du pore de fusion, et le mode de libération du contenu ou de progression vers un pore de fusion complètement ouvert est ralenti.
L'apex d'arginine de C2B aide à contrôler l'ouverture ou la dilatation du pore de fusion et assure la transition entre l'ouverture initiale du pore et un pore de fusion complètement ouvert.
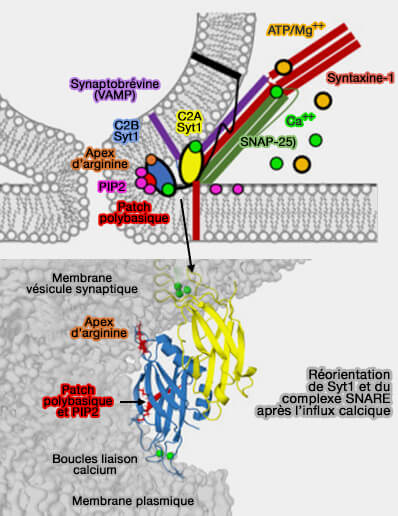
(Figure : vetopsy.fr d'après Nyenhuis et coll)
b. La nouvelle orientation de C2B séquestrerait PI(4,5)P2 ou PIP2 dans la tige membranaire tendue, déstabilisant davantage la membrane et favorisant l'ouverture des pores.
Le patch polybasique est aligné face à une région de courbure négative élevée dans ce modèle.
- Lors de l'interaction avec PIP2, ce lipide serait entraîné dans une interface de courbure négative, qui n'est pas favorable du point de vue énergétique, car, seul, il a tendance à former des micelles.
- La présence de PIP2 dans cette région du pore, entraînée par de fortes interactions électrostatiques avec la face polybasique de C2B, pourrait agir pour déstabiliser un état initial dans le pore de fusion et le conduire à un état complètement ouvert.
Remarques : le domaine C2A, représenté inséré dans la surface de la vésicule, serait l'orientation préférentielle de Syt1 (Phosphatidylinositol 4,5 Bisphosphate Controls the cis and trans Interactions of Synaptotagmin 1 2019).
- Toutefois, le domaine C2A pourrait également être ancré sur la membrane plasmique.
- Les boucles de liaison Ca++ de C2B sont dirigées vers la surface de la membrane plasmique, car C2B a une forte affinité pour les membranes contenant PIP2 à concentration calcique faible, et les boucles sont susceptibles de s'insérer dans la membrane plasmique (PtdInsP2 and PtdSer cooperate to trap synaptotagmin-1 to the plasma membrane in the presence of calcium 2013).
3. À la suite de ce réarrangement lipidique, un pore de fusion s'ouvre et permet au contenu chimique du compartiment donneur d'entrer dans le compartiment accepteur (Molecular mechanism of fusion pore formation driven by the neuronal SNARE complex 2018).
Les étapes de formation du complexe SNARE coïncideraient avec celles de la fusion membranaire.
- L'assemblage des moitiés N-terminales des domaines SNARE induit le contact initial avec la membrane, la formation des tiges de fusion et leur élargissement vers le diaphragme d'hémifusion.
- L'assemblage des moitiés C-terminales des motifs SNARE, mais aussi du lien (linker), fournit l'énergie nécessaire pour compléter la dernière étape de la fusion membranaire, i.e. la formation du pore.
Évolution du pore de fusion
Biologie cellulaire et moléculaireConstituants de la celluleTransport membranaireTransports sans mouvements membranairesTransports passifsTransports actifsTransports avec mouvements membranaires : trafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireCanaux ioniquesTransporteursUniportsPompesCo-transporteursMoteurs moléculairesVoies de signalisation