Les fusions et les fissions spécifiques seront le plus souvent traitées dans des chapitres particuliers aux organites.
Vue d'ensemble
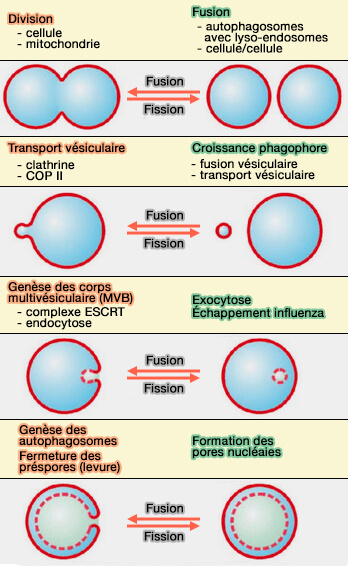
1. Les changements topologiques membranaires se produisent par des événements de fission et de fusion et dans les exemples les plus simples :
la fission se produits lorsqu'une vésicule, i.e. un organite membranaire quelconque, est divisée en deux vésicules,
la fusion se produit lorsque deux vésicules sont fusionnées en une seule.
Plus généralement, un nombre décrivant la topologie de la membrane, appelé caractéristique d'Euler, qui est de 2 lorsqu'on assimile une vésicule à une sphère, est :
soit augmentée de deux lors de la fission, i.e. formation de deux sphères non connectées (2 + 2 = 4),
soit diminuée de deux lors de la fusion (4 - 2 =2).
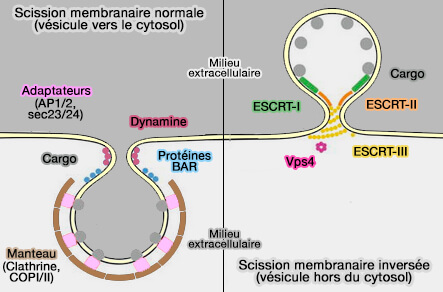
Les changements topologiques externes sont ceux dans lesquels les protéines sont localisées sur le feuillet externe du col membranaire pour être fusionnées, ou brisées par la fission.
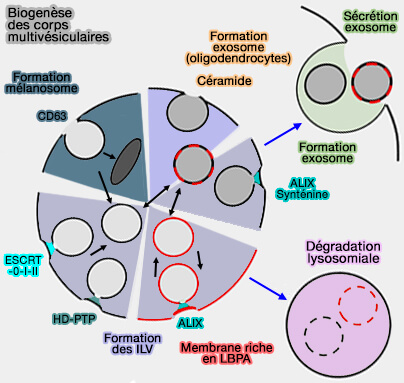
Biogenèse des endosomes ou corps multivésiculaires
(Figure : vetopsy.fr d'après Gruenberg)
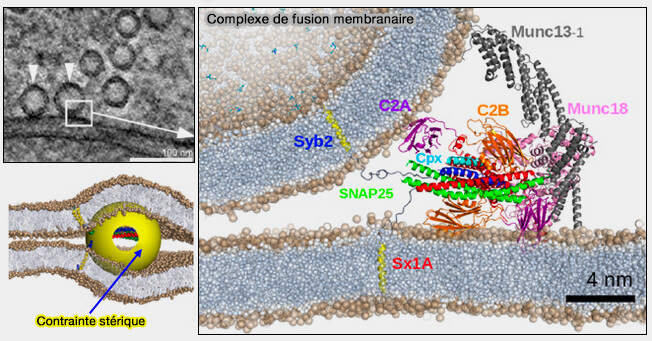
1. La fusion externe est bien documentée (protéines SNARE, protéines virales…).
2. La fission externe peut être intuitivement comprise comme résultant de la constriction du cou par des protéines externes, comme celle bien décrite de la dynamine ( détachement vésiculaire par la dynamine).
Fusion et fission internes
Les changements topologiques internes engagent des protéines se trouvant dans la lumière du cou.
1. La fusion interne peut être comprise intuitivement par l'induction d'une forte courbure locale au site de fusion, comme l'externe par ailleurs.
2. La fission interne est beaucoup moins intuitive.
Dans la fission interne, la constriction des cols membranaires par les machines protéiques présentes dans la lumière devrait être bloquée par les protéines elles-mêmes.
Scission membranaire normale et inversée
(Figure : vetopsy.fr)
Toutes les réactions de fission interne nécessitent l'ESCRT-III qui pourrait être la machinerie générale pour effectuer ce processus.
Le principe général de la réaction de fission interne attribuable à ESCRT-III repose sur un assemblage transitoire sur la membrane pour induire sa courbure, qui n'est pas encore bien comprise, qui permettrait de resserrer le col membranaire potentiellement important au point où la fission se produit.
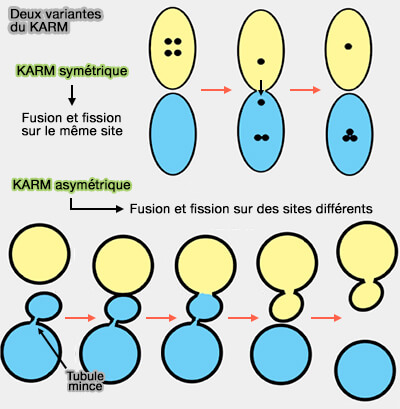
b. La variante asymétrique du KARM présuppose que la fusion/fission peut se produire sur des sites différents, appelé aussi modèle de maturation des transporteurs (CMM ou carrier-maturation model).
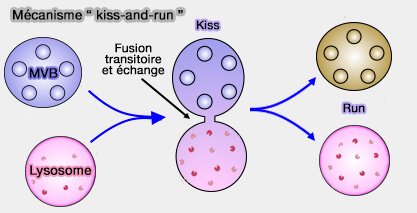
En accélérant le recyclage des vésicules, le " kiss-and-run " fournit un mécanisme efficace pour faciliter la libération prolongée.
Le modèle " kiss-and-run " (KARM) est de plus en plus incriminé dans le trafic vésiculaire que ce soit dans l'endocytose rapide que dans la voie sécrétoire ( KARM de l'EGT et KARM de l'IGT).
En outre, des protéines qui ont des rôles contradictoires dans la fusion et la fission des lysosomes, comme Lyst1 (LYSososmal-Trafficking regulator) pourrait former des échafaudages qui permettent une fusion partielle et coordonner la fission
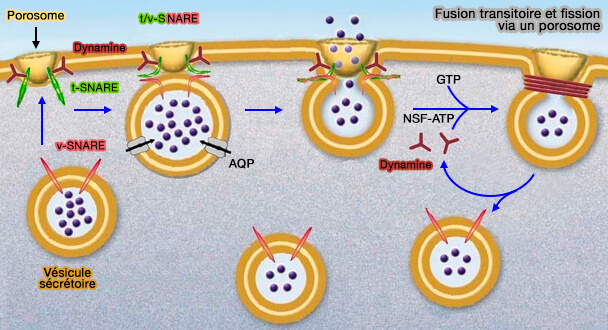
Fusion transitoire et fission via un porosome
(Figure : vetopsy.fr d'après Jena)
On pourrait imaginer un échafaudage d'un complexe semblable au porosome et Lyst1, une structure en coupe qui serait intégrée dans le membrane plasmatique et dans laquelle une vésicule exocytaire s'arrimerait pour former un tunnel à travers lequel s'écoulerait le contenu.






 détachement vésiculaire par la dynamine).
détachement vésiculaire par la dynamine).