La fusion de deux vésicules lipidiques entraîne des changements dans la topologie membranaire et la conformation lipidique.
Les membranes doivent être mises en contact direct, ce qui nécessite le retrait des coques d'hydratation des feuillets externes.
Pour contourner cet handicap énergétique, des machineries se sont mises en place pour faciliter la fusion et une question ouverte importante est de savoir si elles agissent :
soit en surmontant les barrières d'énergie libre, ce qui paraît être le cas des complexes SNARE et les protéines de fusion virale,
soit, par le rôle catalytique, en abaissant ces barrières.
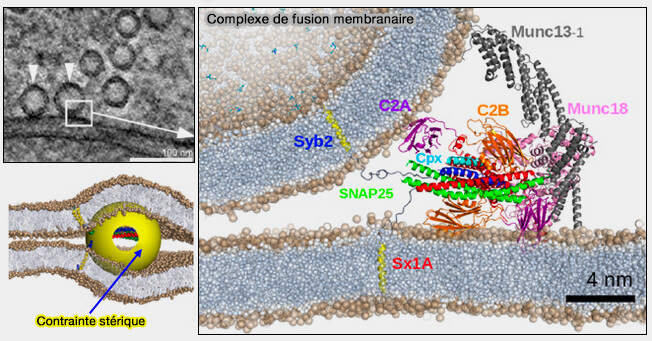
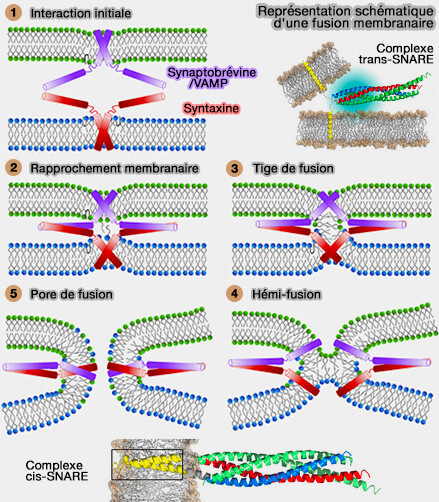
Complexe de fusion membranaire
(Figure : vetopsy.fr d'après Grubmüller et coll)
2. La force mécanique exercée par les protéines de fusion peut permettre l'apposition des deux membranes, au moins pour induire des contacts ponctuels ( rapprochement membranaires lors de la fusion).
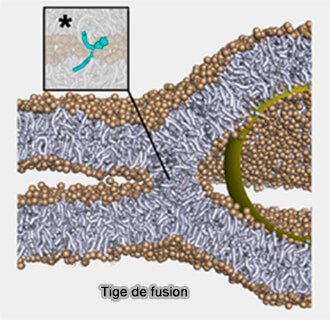
a. Les protéines de fusion sont censées déformer une petite zone de membrane pour former des structures en forme de pointes qui font saillie vers le partenaire de fusion et favorisent l'étalement des lipides du feuillet externe, i.e. formation d'une tige de fusion.
Tige de fusion (hémifusion)
(Figure : vetopsy.fr d'après Grubmüller et coll)
Ce processus favorise la fusion des feuillets externes, conduisant à un diaphragme d'hémifusion dans lequel les lipides peuvent passer entre les feuillets externes, mais les feuillets internes séparés empêche le mélange des contenus.
L'hémifusion semble être un intermédiaire universel qui peut être observé dans les réactions de fusion entre bicouches purement lipidiques ainsi que dans la fusion protéique entre membranes physiologiques.
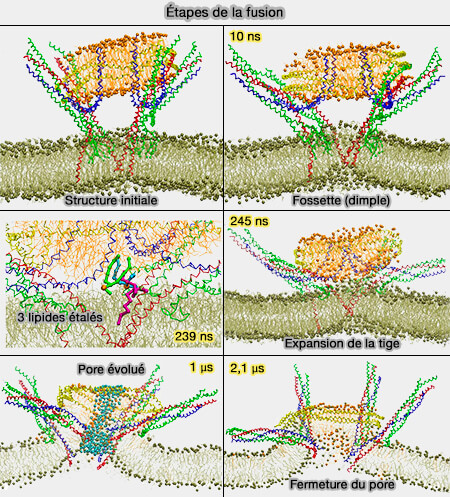
b. La fusion complète nécessite que les feuillets internes fusionnent pour former un pore de fusion initial qui se trouve dans un état d'énergie minimale locale, i.e. il est métastable.
Les feuillets internes doivent se rapprocher, ce qui nécessite que la tige se comprime et s'élargisse ( élargissement de la tige).
La force mécanique fournie par les protéines SNARE est supposé entraîner une déformation locale des feuillets internes dans la structure d'hémifusion, imposer des réarrangements lipidiques et la génération d'un pore de fusion initial.
Étapes de la fusion
(Figure : vetopsy.fr d'après Sharma et coll)
Comment les protéines de fusion arrivent elles à leur fin est un enjeu majeur et l'un des problèmes centraux à résoudre à l'heure actuelle.
1. En effet, par souci de simplicité et de clarté, les résultats des études in vitro et in vivo sont généralement interprétés en supposant que les SNARE sont les seules forces motrices de la fusion, i.e. en faisant abstraction des interactions des SNARE avec d'autres protéines :
2. Toutefois, le développement de nouveaux outils a fait grandement avancé la compréhension de la fusion membranaire.
a. Tout d'abord, le développement de processeurs plus rapides, et en particulier d'accélérateurs graphiques (GPU), ont facilité l'échantillonnage informatique de l'espace des phases.
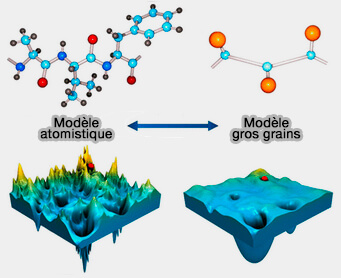
Modèle atomistique versus gros grains
(Figure : vetopsy.fr d'après Kmiecik et coll)
b. D'autre part, du côté de la modélisation moléculaire, les modèles dits gros grains comme, par exemple, le modèle de Martini) cartographient des groupes chimiques individuels en un seul site d'interaction, i.e. ils sont 200 fois plus précis que les simulations atomistiques (Coarse-Grained Protein Models and Their Applications 2016).
La modélisation gros grain consiste à considérer des groupes d’atomes qui interagiront les uns avec les autres comme les atomes dans les approches atomistiques.
Entrons maintenant un peu plus dans le détail, en gardant bien à l'esprit que la fusion est un processus continu dont toutes les étapes ne sont pas connues avec précision !





 machinerie de fusion).
machinerie de fusion).