


Fusion membranaire : mécanisme
Évolution du pore de fusion
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Transport membranaire
- Transports membranaires sans mouvements membranaires
- Transports membranaires avec mouvements membranaires : trafic vésiculaire
- Protéines membranaires
- Moteurs moléculaires
- Voies de signalisation

Les pores de fusion dépendants des SNARE sont généralement considérés comme des structures transitoires.
Les pores de fusion sont métastables, mais ils peuvent osciller (pore flickering) entre une fusion complète ou un retour à une hémifusion.
Expansion du pore de fusion
1. Les pores de fusion dépendants des SNARE sont généralement considérés comme des structures transitoires, avec des durées de vie de la gamme inférieure à la seconde (Fusion pores and their control of neurotransmitter and hormone release 2018 et Dynamics and number of trans-SNARE complexes determine nascent fusion pore properties 2018).
L'expansion du pore de fusion s'oppose :
- à l'énergie libre liée à l'extension de son périmètre membranaire courbe, i.e. Il faudrait la présence d'une force extérieure, comme la présence de pression osmotique ou une couche de protéines en croissance sur le col du pore de fusion,
- à la création d'une zone membranaire en excès lors de l'expansion des pores (croissance du volume de la vésicule).
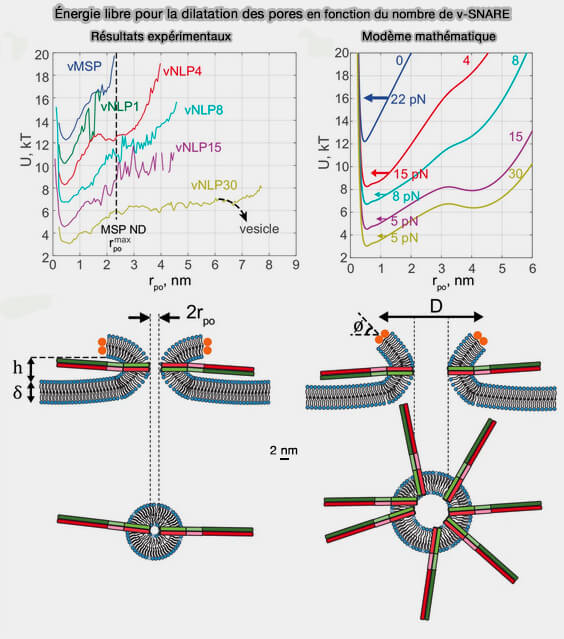
(Figure : vetopsy.fr d'après Wu et coll)
2. L'expansion du pore pourrait être favorisée par l'incorporation d'un nombre croissant de complexes SNARE dans la membrane entourant le pore et qui augmente progressivement la conductance du pore de fusion (pour une définition de la conductance, Fusion pores and their control of neurotransmitter and hormone release 2018).
Les répulsions stériques entre le nombre croissant de complexes SNARE peuvent renforcer l'élargissement du pore (Dilation of fusion pores by crowding of SNARE proteins 2017).
Pores quiescents ou nanopores
Vue d'ensemble
1. Chez la levure, les vacuoles sont reliés par des pores de fusion nanoscopiques pendant plusieurs minutes (SNARE-mediated membrane fusion arrests at pore expansion to regulate the volume of an organelle 2018).
- Ces pores sont très étroits, i.e. diamètre de 1 nm ou moins, et peuvent rester quiescents pendant une longue période.
- Leur existence ne peut être démontrée par des marqueurs lipidiques fluorescents, d'où leur appellation ironique de " trous noirs " (SNAREs, tethers and SM proteins: how to overcome the final barriers to membrane fusion? 2020).
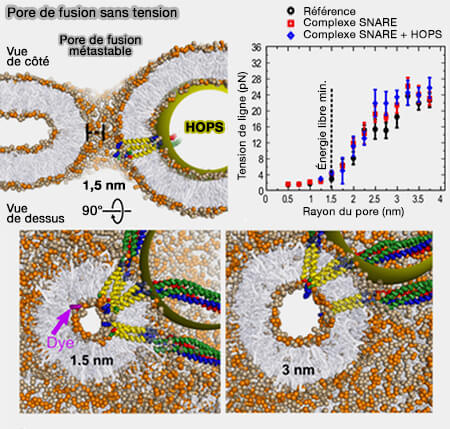
(Figure : vetopsy.fr d'après D'agostino et coll)
2. Toutefois, ces petits pores permettaient toujours le passage de domaines transmembranaires marqués malgré leurs extrémités C-terminales chargées, illustrant ainsi une capacité restante à conduire les ions (SNARE-mediated membrane fusion arrests at pore expansion to regulate the volume of an organelle 2018).

Existe-t-il des nanopores relativement stables dans d'autres étapes de fusion physiologique impliquant les SNARE dans lesquelles ils seraient en conformation d'hémifusion (Hemifusion in Synaptic Vesicle Cycle 2017) ?
Mécanismes possibles
Les pores quiescents limitent la diffusion à travers le pore en réduisant la dynamique de l'eau par encombrement moléculaire dans le petit pore (Water Dynamics in the Hydration Shells of Biomolecules 2017).
- La fermeture des pores augmente la contrainte de flexion sur les bicouches (Teardrop shapes minimize bending energy of fusion pores connecting planar bilayers 2013).
- Elle coïncide également avec la déshydratation défavorable des groupes de tête lipidiques qui est nécessaire pour surmonter la répulsion d'hydratation entre les feuillets luminales (Interbilayer repulsion forces between tension-free lipid bilayers from simulation 2013).
1. À des tailles de pores <1 nm, les répulsions d'hydratation domineraient l'énergie libre du pore. Cela suggère que le pore de fusion in vivo doit être soumis à une force de " compression " supplémentaire qui compense la répulsion de l'hydratation luminale dont la nature est floue à l'heure actuelle.
a. Ce peut être la présence d'attractions de nature électrostatique dans la lumière du pore.
- Elles pourraient se produire entre les groupes de tête lipidiques chargés, les résidus chargés dans les extrémités C-terminales luminales des SNARE et les cations divalents (Fusion pores and their control of neurotransmitter and hormone release 2017).
- En outre, elles inhiberaient la formation du diaphragme d'hémifusion car l'hémifission diminuerait le nombre de têtes désormais favorables interactions de groupe à l'intérieur du pore.
b. Si on augmente de 5% les lipides des feuillets externes, cette asymétrie peut faciliter les pores de taille inférieure au nanomètre, en particulier lorsque les lipides de type PE (phosphatidyléthanolamine) tapissent en outre l'intérieur du pore.
Les vacuoles reliant les pores de fusion nanoscopiques stables pourraient donc s'expliquer par des différences de densité lipidique des feuillets et de leur composition en conséquence de l'amarrage vacuole-vacuole.
2. Un pore tapissé de plusieurs complexes SNARE affiche une plus grande stabilité contre l'hémifission, conformément aux observations expérimentales.
a. Un pore qui se ferme expulse activement les SNARE et les plient pour générer une contrainte.
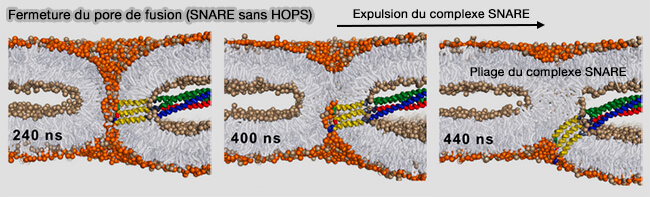
(Figure : vetopsy.fr d'après D'agostino et coll)
b. Ce mouvement devrait être entravé par la rigidité et la longueur du complexe SNARE, ainsi que par son attachement au complexe HOPS, qui est ancré dans les deux membranes.
La répulsion stérique que HOPS impose au site de fusion réduit la courbure effective de l'intérieur du pore, ce qui, par analogie avec le rôle de la courbure dans la fusion membranaire, stabilise le pore en s'opposant à la formation d'une tige d'hémifission.

Un seul complexe SNARE avec HOPS réduit la probabilité de fermeture des pores jusqu'à 80%.
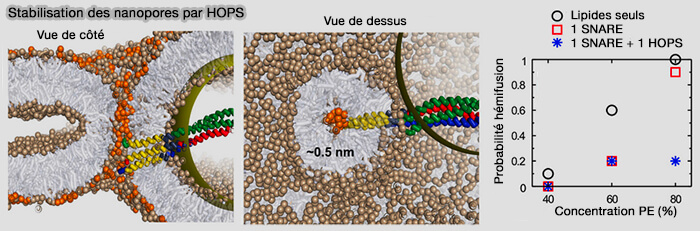
(Figure : vetopsy.fr d'après D'agostino et coll)
3. Toutefois, il semble que la présence de zones de contact vésicule-vésicule étroitement ancrées restent métastables, même après clivage enzymatique des complexes SNARE (SNARE and regulatory proteins induce localmembrane protrusions to prime docked vesicles forfast calcium-triggered fusion 2014).
a. Cette attraction serait d'origine osmotique en raison de l'accès stériquement restreint des ions à l'espace étroit entre les bicouches lipidiques apposées chargées (Electrostatic double-layer interaction between stacked charged bilayers 2017).
- Ce contact n'est pas observée en présence d'un intermédiaire d'hémifusion formé ou d'un pore de fusion soulève la question de savoir comment ces états ancrés sont mobilisés pour la fusion.
- Les bicouches empilés sont séparées de 1,1 et 1,3 nm, distance supérieure à celle la fusion membranaire qui nécessite une distance inférieure à 1 nm afin de former un pont lipidique crucial.
- L'adhérence s'oppose très probablement à la fois à la formation et à l'expansion du pore de fusion ainsi qu'à la formation d'intermédiaires d'hémifusion, car cela réduirait la quantité de surface de contact membrane-membrane favorable.
b. L'adhérence pourrait aider à surmonter les affrontements stériques et aider à la fois la fermeture éclair et la structuration du complexe SNARE en fournissant une force motrice supplémentaire et peut-être essentielle pour rapprocher suffisamment les membranes.
Petites versus grandes vésicules
Il n'est pas évident que les paramètres pertinents pour une fusion efficace se chevauchent entièrement entre les petites vésicules exocytaires, qui sont optimisées pour une fusion rapide et temporellement bien contrôlée, et vésicules plus grandes ou les organites, dans lesquels les barrières énergétiques et les forces d'indentation concomitantes devraient être beaucoup plus grande.
1. Les courbures membranaires sont différentes.
- Les sites de fusion entre deux très grandes vésicules peuvent être considérés approximativement comme plats, ce qui laisse peu de place aux mouvements important des SNARE et de leurs protéines associées perpendiculairement aux membranes.
- Les sites de fusion entre de très petites vésicules, i.e. vésicules synaptiques de diamètre <40 nm, possèdent de très grandes courbures membranaires et offrent beaucoup plus d'espace pour que les complexe SNARE se réorientent eux-mêmes.
Cela peut être un facteur important réduisant la stabilité des pores de fusion car cela peut permettre au complexes SNARE de diffuser plus facilement hors du pore de fusion.
2. Une autre différence pertinente concerne la taille de la zone de contact.
a. Cette zone est très grande entre les organites amarrés, i.e. plusieurs μm2 pour les vacuoles de levure.
- Le décalage des complexes SNARE volumineux peuvent imposer la formation de zones de contact plus larges, qui accumulent plus de tension de membrane, accélèrent la croissance de la zone de contact et permettent le tension générée pour favoriser la fusion avant qu'elle ne puisse se détendre.
- L'asymétrie des pores concomitante est due à l'abaissement de énergie de flexion de la membrane entre le pore de fusion et le bord incurvé membranaire qui lui est associé, i.e. la présence de complexes volumineux comme HOPS peut augmenter l'énergie libre interface du vertex, en augmentant l'angle de contact entre les vacuoles et en imposant un effet stérique direct sur les membranes.
b. Les vésicules synaptiques entrent en contact au travers de surface très faible, i.e. ponctuelles (The Morphological and Molecular Nature of Synaptic Vesicle Priming at Presynaptic Active Zones 2014). Pour de si petites vésicules, la taille du pore de fusion précoce occupe déjà une fraction significative d'une zone d'adhérence potentiellement présente, ce qui est bénéfique.
- De plus, la minimisation de la contrainte de courbure grâce à la fusion de petits vésicules peuvent entraîner l'expansion du pore de fusion car leur contrainte de courbure peut être importante et compétitive avec la tension de ligne du pore de fusion.
- Ainsi, de très petites vésicules devraient être prédisposées pour une progression rapide vers l'ouverture et l'expansion des pores de fusion.
Retour à la fusion membranaire
Biologie cellulaire et moléculaireConstituants de la celluleTransport membranaireTransports sans mouvements membranairesTransports passifsTransports actifsTransports avec mouvements membranaires : trafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireCanaux ioniquesTransporteursUniportsPompesCo-transporteursMoteurs moléculairesVoies de signalisation