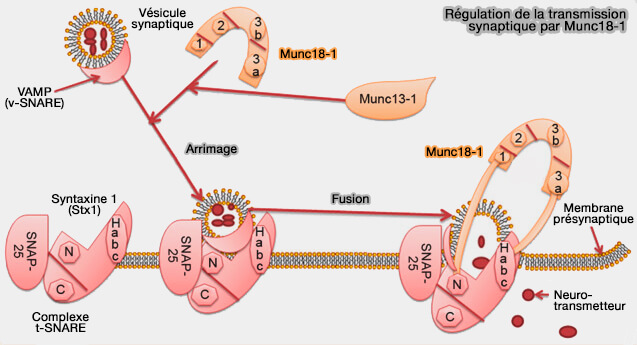
Régulation de la transmission synaptique par Munc18-1
(Figure : vetopsy.fr d'après Tang et coll)
Chaque protéine SM interagit avec une SNARE ou plusieurs complexes de SNARE (SNAREs and traffic 2005).
Les mécanismes moléculaires sous-jacents à la fonction SM sont difficiles à déchiffrer, car les interactions entre les protéines SM et les SNARE apparentées sont très variées.
Complexe Munc18-Qa
Vue d'ensemble
1. Munc18-1 régule la transmission des neurotransmetteurs en favorisant ou en inhibant l'assemblage du complexe SNARE principalement en se combinant avec la syntaxine-1 (Stx1), Qa-SNARE.
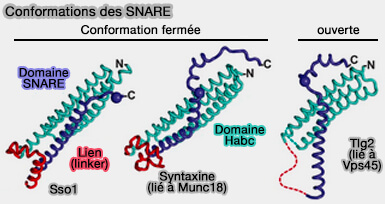
Conformations des SNARE
(Figure : vetopsy.fr d'après Eisenmann et coll)
Le motif SNARE, composé par une seule hélice dans le complexe cis-SNARE, montre plusieurs hélices plus courtes et son extrémité C-terminale est désordonnée.
La plupart de ses résidus centraux sont séquestrés lors d'interactions avec le domaine Habc (couches -7 à +1) ou Munc18-1 (couches +2 et +3), empêchant la syntaxine-1 de s'associer à d'autres SNARE.
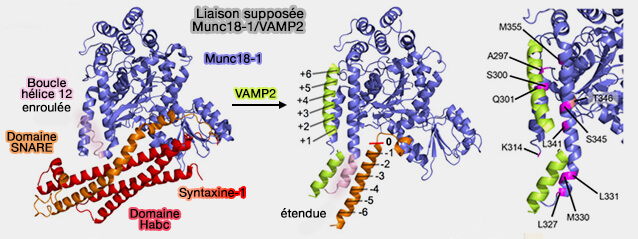
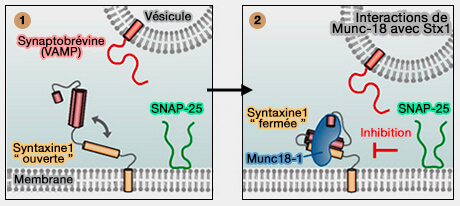
Modèle d’ouverture de la syntaxine 1 en complexe avec Munc18
(Figure : vetopsy.fr d'après Stepien et coll)
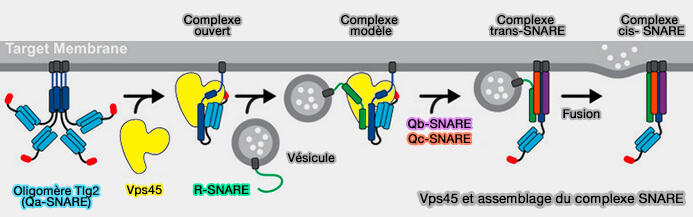
En général, les protéines SM protègent les intermédiaires appropriés d'assemblage du complexe SNARE de NSF/α-SNAP, tout en permettant à NSF/α-SNAP de recycler les produits des réactions de désassemblage SNARE, ce qui a été démontré pour Munc18-1, Vps33 et Sly1 ( assemblages impropres).
Munc18-1 combiné à Stx1 de conformation " fermée " sert de modèle pour l'assemblage SNARE ( modèle de l'assemblage SNARE).
Remarque : le domaine central MUN de Munc13-1 se liera avec la syntaxine-1 pour libérer le domaine Hc du complexe Munc18/syntaxine et pour former le complexe ternaire syntaxine-1/SNAP-25/VAMP2 qui favorise la fusion des vésicules synaptiques ( Munc-13 et complexe SNARE).
Protéines SM et conformations " fermées "
Est-ce que les protéines SM piègent toujours les Qa-SNARE dans des conformations fermées ?
Dans la plupart des articles plus tardifs que 2008, il manquait toujours un motif aux structures proposées.
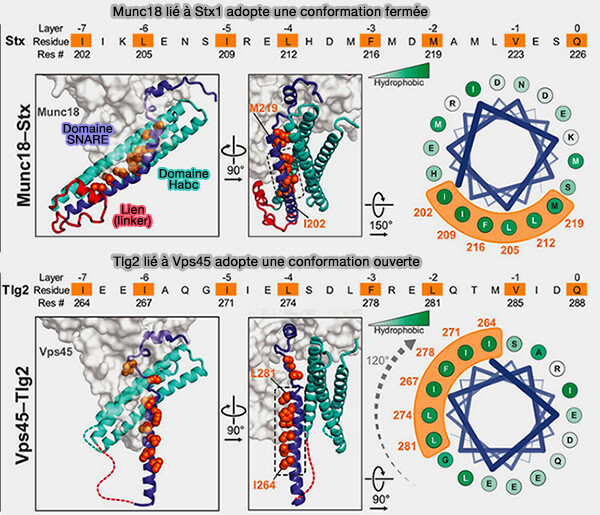
Comparaison Munc18/Stx1 et Vps45/Tlg2
(Figure : vetopsy.fr d'après Eisenmann et coll)
Bien que Vps45 engage les mêmes régions Qa-SNARE que Munc18-1, i.e. le peptide N-terminal, le domaine Habc et le domaine SNARE, la conformation du SNARE lié est beaucoup plus ouverte.
Le motif SNARE interagit de la même manière avec la protéine SM, centrée autour des couches +2 et +3, mais interagit assez peu avec le domaine Habc.
Vps45 et assemblage du complexe SNARE
(Figure : vetopsy.fr d'après Eisenmann et coll)
Inversement, l'épingle à cheveux hélicoïdale adopte une conformation enroulée dans la structure Munc18-1-Stx1, avec la pointe de l'épingle à cheveux repliée qui enfouit une partie substantielle du site de liaison R-SNARE.
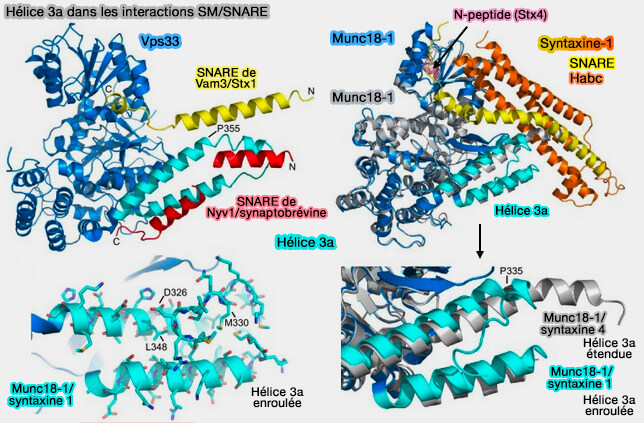
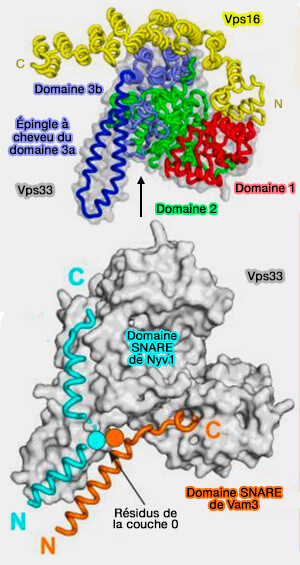
Complexe Vps33/Nyv1
(Figure : vetopsy.fr d'après Baker et coll)
Les NTD des domaines SNARE sont hélicoïdaux et correctement alignés pour l'assemblage du complexe SNARE.
Les CTD, en revanche, divergent, le R-SNARE suivant la rainure dans l'épingle à cheveux hélicoïdale, tandis que le Qa-SNARE suit le même chemin que dans la structure fermée Munc18-1-syntaxine-1.
Ce modèle est évocateur d'un complexe SNARE semi-zippé.
La protéine SM fonctionnerait comme une matrice, nucléant l'assemblage SNARE en maintenant les NTD des motifs R- et Qa-SNARE alignés pour qu'ils puissent se lier aux NTD des Qb- et
Qc-SNARE.
La formation de ce complexe serait une étape limitante pour la fusion membranaire médiée par SNARE in vitro et in vivo.
La majorité des motifs R- et Qa-SNARE contribuent à la stabilité du complexe de modèles :
La suppression du domaine Habc inhibe la formation du complexe matrice.
La région de liaison, i.e. Habc et épingle à cheveux des SM qui est bien ordonnée dans le complexe Munc18-1-syntaxine-1, est susceptible d'être dépliée dans le complexe matrice.
De nombreuses questions restent en suspens, et les modèles évoluent, surtout qu'une nouvelle protéine, Munc13-1, entre en jeu. Le modèle de chaperonnage de Munc18-1/Munc13-1 est étudié dans un chapitre spécial.





.jpg)
 assemblages impropres).
assemblages impropres).