Avec les synaptotagmines, elles ne sont pas nécessaires à la fusion en soi, mais ils permettent l'accumulation, au niveau de la membrane plasmique, d'une réserve (pool) de vésicules prêtes à fusionner en attente du signal, i.e. entrée de calcium pour une libération synchrone.
1. Quatre gènes de complexine différents, cplxI–cplxIV ont été décrits chez la souris, et des orthologues correspondants existent également dans le génome humain.
Les sous-groupes diffèrent principalement par leur domaine C-terminal qui, dans le cas de Cplx3/4 porte une extension avec un motif de boîte CAAX pour l'isoprénylation, i.e. ajout d'un groupe isoprénoïde au résidu cystéine à son extrémité C-terminale.
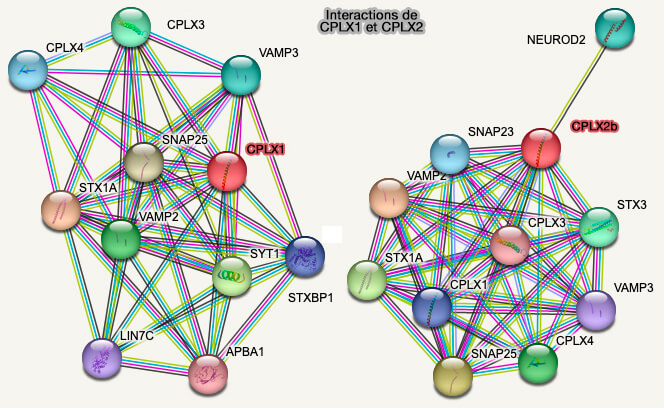
Interactions de CPLX1 et CPLX2
(Figure : vetopsy.fr d'après String)
2. Les quatre isoformes de la complexine sont principalement exprimées dans le système nerveux central, mais Cplx4 est surtout retrouvée dans les synapses à ruban (ribbon synapse) de la rétine.
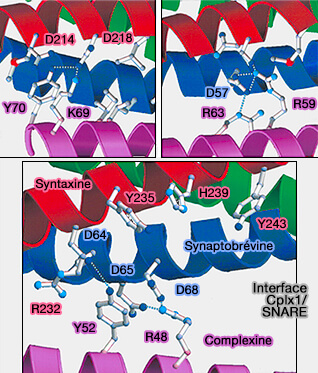
Interactions hélice CH/SNARE
(Figure : vetopsy.fr d'après Chen et coll)aux résidus 214 à 232 de la syntaxine.
aux résidus 47 à 68 de la synaptobrévine.
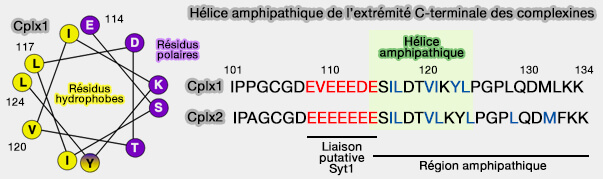
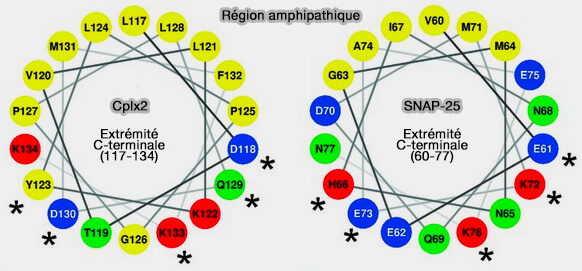
La liaison à la complexine recouvre la couche polaire centrale (couche 0) du complexe SNARE.
Deux résidus tyrosine (Y52 et Y70) et trois résidus arginine (R48, R59 et R63) de la complexine semblent être essentiels pour la liaison au complexe SNARE.
La chaîne latérale de Y52 établit des contacts étendus avec la synaptobrévine 2/VAMP2, y compris une liaison hydrogène avec la chaîne latérale D64.
Y70 est impliqué dans des contacts étendus avec la syntaxine, formant une liaison hydrogène avec D218.
Les trois résidus arginine de la complexine forment des ponts salins avec trois résidus aspartate de VAMP2 (D57, D65 et D68) et sont également impliqués dans les interactions hydrophobes.
Les résidus M62, I66, L222 et M229 de Stx1, V50 et L54 de VAMP2 participent à des interactions hydrophobes supplémentaires, tandis qu'un résidu de lysine de la complexine (K69) forme un autre pont salin avec un résidu d'aspartate de la syntaxine (D218).
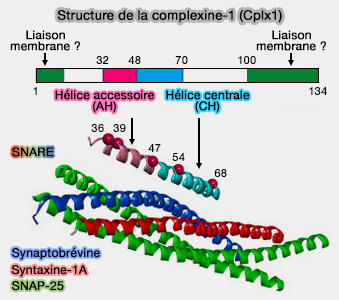
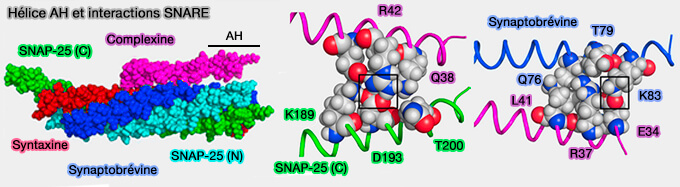
Bien que ce motif ne soit pas essentiel pour la liaison SNARE, les résidus N-terminaux (acides aminés 41 à 47) semblent améliorer la liaison SNARE de l'hélice centrale.
La formation de l'hélice semble commencer dans l'hélice accessoire et se propage ensuite dans la région de l'hélice centrale, stabilisant ainsi potentiellement l'hélice centrale et augmentant la liaison SNARE.
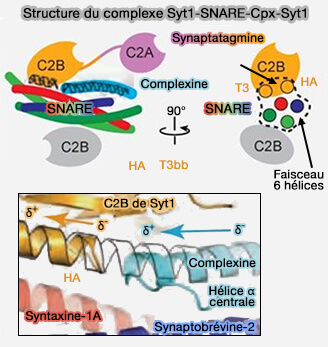
Complexe Syt1-SNARE-Cplx-Syt1
(Figure : vetopsy.fr d'après Zhou et coll)




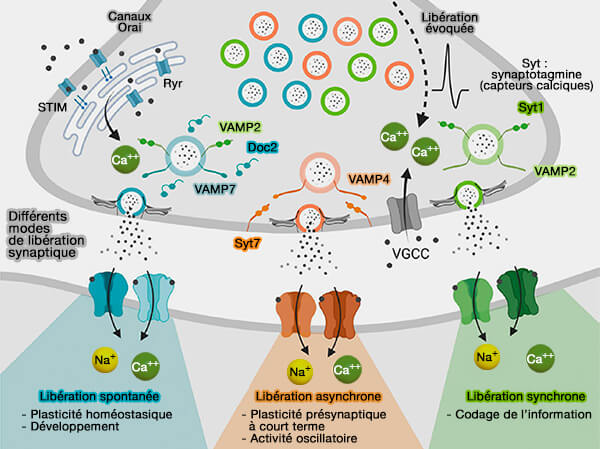

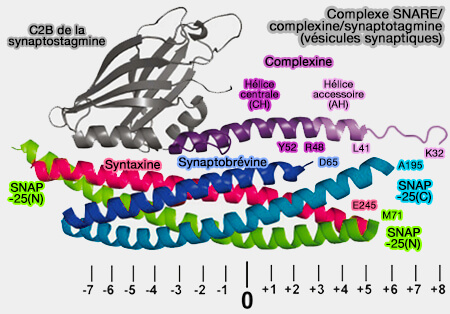
 modèles de régulation des SNARE par la complexine/synaptotagmine).
modèles de régulation des SNARE par la complexine/synaptotagmine).