dans les cellules musculaires (striées ou lisses), et jouent alors un rôle essentiel dans la contraction des muscles (squelettiques, cardiaque et lisses),
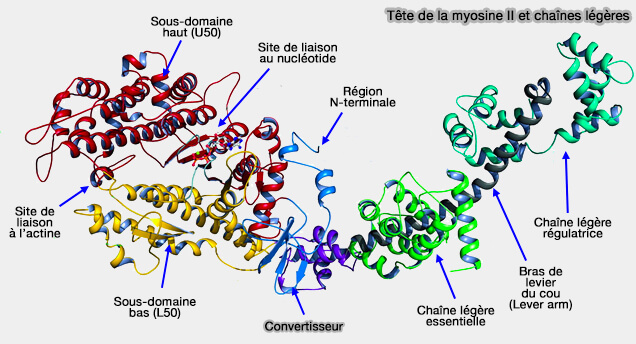
C'est au niveau de la tête de myosine (tête S1) que se situe l'activité ATPasique et le site de liaison à l'actine qui augmente fortement cette activité.
Domaine moteur de la tête
Structure générale
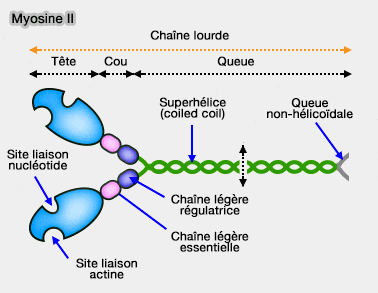
Structure de la myosine II
(Figure : vetopsy.fr)
Le domaine moteur est inséré dans la partie globulaire (d'environ 780 acides aminés) de la tête de la myosine, région N-terminale de la chaîne lourde, qui contient aussi le site de liaison à l'actine.
Ce moteur contient des hélices-α disposées autour d'un coeur, domaine constitué d'un feuillet β (β sheet) à 7 brins, appelé transducteur.
Si ce moteur est bien conservé par l'évolution, sa régulation par des liaisons ioniques ou des modifications post-traductionnelles est très largement différenciée suivant les différentes isoformes (Principles of Unconventional Myosin Function and Targeting 2011).
Sous-domaines du domaine moteur
Le domaine moteur peut être subdivisé en 4 sous-domaines.
1. Le sous-domaine N-terminal.
2 et 3. Les sous-domaines de 50 kDa, haut (U50 : U pour " Upper ") et bas (L50 : L pour " Lower ") peuvent être considérés comme les pinces d'une tenaille : le déplacement de U50 provoque la fermeture de la tenaille pour " enserrer " l'actine.
Une large fissure (poche nucléotidique) entre U50 et L50 contient 3 structures de liaison nucléotidique :
une P-loop (Phosphate binding loop : séquence riche en glycine suivie d’une lysine conservée puis d’une sérine ou une thréonine, présente dans la majorité des ATPases et des GTPases) qui se lie à l'ATP,
On retrouve quelquefois un domaine SH3 qui n'existe pas dans certaines myosines (myosine I) et qui n'est pas impliqué dans l'activité motrice.
Cou de la myosine II
Le cou est composé de deux domaines (ou motifs) IQ (calmodulin-binding motif), motifs de 25 acides aminés ([I,L,V]QxxxRGxxx[R,K]), formant une hélice-α amphiphylique.
1. Les domaines IQ se lient à la calmoduline et aux chaînes légères calmoduline-like.
Structure de la myosine
(Photographie : greatcourse.cnu.edu.cn)
La chaîne légère essentielle (ELC la plus proche du domaine moteur) de la myosine II du muscle lisse liée au premier motif IQ est en contact avec une boucle du domaine moteur qui module la libération d'ADP.
: chaîne légère essentielle (ELC la plus proche du domaine moteur) et la chaîne légère régulatrice.
La la chaîne légère régulatrice (RLC), lié au second motif IQ, est régulée par phosphorylation d'un site de de son extrémité N-terminale.
2. Le cou, formés par les motifs IQ et les chaînes légères, agit comme un bras de levier qui amplifie le mouvement généré par le domaine de moteur (Fifty Ways to Love Your Lever: Myosin Motors 1996).
Le nombre de motifs IQ détermine la longueur du bras de levier et par conséquent la taille de pas du moteur de myosine.
Cependant, les motifs IQ et les chaînes légères associées ne sont pas simplement des dispositifs mécaniques passifs, mais influencent également les propriétés chimio-mécaniques.
Cette superhélice est chargée négativement sur une grande partie de sa structure. Toutefois, elle est formée de séquence de 28 acides aminés alternativement chargés négativement et positivement qui maintiennent un alignement parfait entre les molécules de myosine.
La courte extrémité C-terminale non-hélicoïdale de 47 acides aminés a un rôle est essentiel pour la bonne taille des filaments et pour éviter les inconvénients inhérents à une queue spiralée.
les filaments épais dans les cellules musculaires striées en relation avec les filaments fins pour former les myofibrilles et provoquer, par leur interaction, la contraction (mécanisme de la liaison mysoine II-actine),
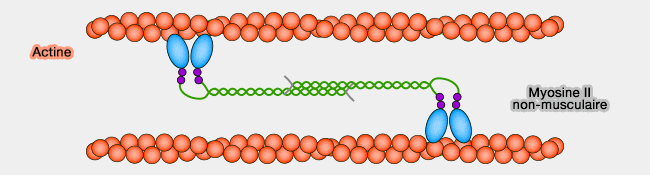
les faisceaux antiparallèles dans les cellules non-musculaires.
Liaison entre myosine II non-musculaire et actine
(Figure : vetopsy.fr)