


Fusion membranaire : mécanisme
Formation de la tige de fusion (2)
Rôles des lipides et de la synaptotagmine
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Transport membranaire
- Transports membranaires sans mouvements membranaires
- Transports membranaires avec mouvements membranaires : trafic vésiculaire
- Protéines membranaires
- Moteurs moléculaires
- Voies de signalisation

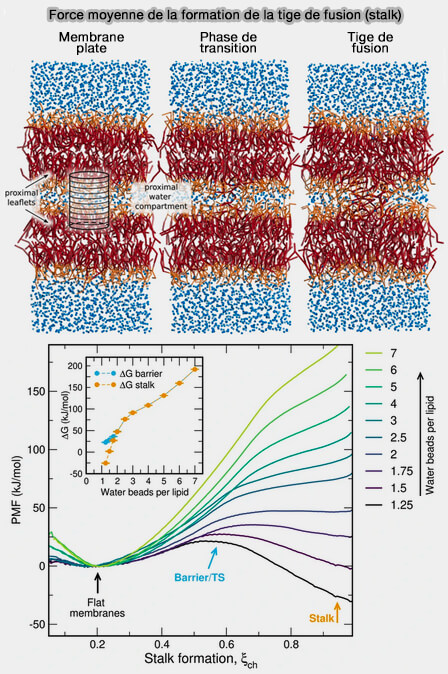
La forme des lipides joue un rôle crucial pour surmonter la répulsion d'hydratation entre deux membranes et diminue le travail total requis pour former une tige.
Lipides membranaires et tige de fusion
(Figure : vetopsy.fr d'après Poojari et coll)

Pour plus de précisions, vous pouvez lire : Free energies of membrane stalk formation from a lipidomics perspective (2021).
Vue d'ensemble
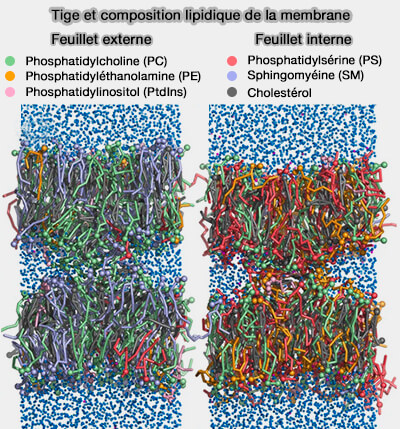
Les membranes cellulaires eucaryotes contiennent plus de dix classes de lipides, tandis que chaque classe comprend des centaines d'espèces chimiques différentes (Membrane lipids: where they are and how they behave 2008 et Lipidomics: coming to grips with lipid diversity 2010).
1. La complexité des membranes est encore augmentée par l'asymétrie membranaire, c'est-à-dire par des compositions lipidiques distinctes dans les deux feuillets (Asymetric Lpid Membranes: Towards More Realistic Model Systems 2015).
Par exemple, dans la membrane plasmique :
- le feuillet externe est enrichi en sphingomyéline (SM) et en phosphatidylcholine (PC),
- le feuillet interne comprend plus de phosphatidyléthanolamine (PE), de phosphatidylsérine (PS) et de phosphatidylinositol (PtdIns).
(Figure : vetopsy.fr d'après Poojari et coll)
2. En outre, le degré de polyinsaturation des lipides dans le feuillet interne est environ le double de celui du feuillet externe (Plasma membranes are asymmetric in lipid unsaturation, packing, and protein shape 2020).
Courbure des lipides
Étant donné que les structures intermédiaires le long de la voie de fusion impliquent des membranes très incurvées, la courbure intrinsèque des lipides influence la cinétique de fusion.
1. Par exemple, lorsqu'elle est ajoutée aux feuillets externes, la lysophosphatidylcholine (LPC) en forme de cône inversé inhibe la formation de la tige car elle est incompatible avec la grande courbure négative au bord de la tige.
2. En revanche, les lipides PE (phosphatidyléthanolamine) insaturés en forme de cône ou le diacylglycérol favorisent la formation de tiges.
- Les acides gras insaturés rendent les lipides plus coniques car les doubles liaisons augmentent le désordre de la queue et, par conséquent, le volume effectif des queues lipidiques.
- En plus de ces effets par la forme géométrique des lipides, l'abondance accrue de doubles liaisons augmente la flexibilité conformationnelle des lipides, permettant ainsi à la membrane de s'adapter plus facilement à la courbure, ce qui peut favoriser davantage la fusion (From zero to six double bonds: phospholipid unsaturation and organelle function 2015).
3. Si la fusion se produit entre deux membranes déjà très incurvées, les lipides " fusiogènes " à courbure spontanée négative, par exemple, le POPE (1-palmitoyl-2-oléoyl-phosphatidyléthanolamine) et le cholestérol ne devraient pas abaisser davantage la barrière d'expansion de la tige (Expansion of the fusion stalk and its implication for biological membrane fusion 2014), contrairement à la réaction de fusion à faible courbure membranaire (A tethering complex drives the terminal stage of SNARE-dependent membrane fusion 2017).
L'hydratation des groupes de tête lipidiques à la circonférence de la fosette de la tige (dimple stalk) est en fait plus grande que celle du lipide correspondant dans une membrane plate entièrement hydratée, i.e. la tige n'est effectivement pas une structure incurvée négativement par rapport à une membrane plane.
(Figure : vetopsy.fr d'après Risselada et coll)
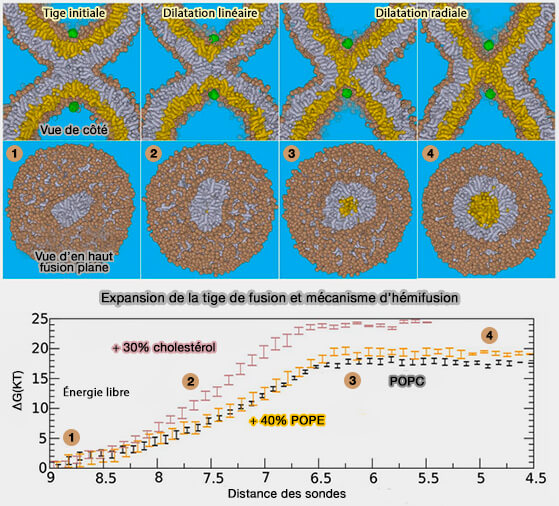
L'élargissement de la tige dans la fusion des fossettes (courbure élevée de la membrane) ne semble donc pas limité par une déshydratation coûteuse des groupes de tête lipidiques, mais plutôt par un étirement relatif des feuillets internes pour former un contact initial.
- Cela explique pourquoi le cholestérol peut s'opposer à l'élargissement de la tige dans la fusion des fossettes puisqu'il augmente les modules élastiques des feuillets.
- En revanche, la présence de lipides insaturés devrait donc favoriser l'élargissement de la tige puisque ces lipides réduisent les modules élastiques.
C'est peut-être l'une des nombreuses raisons pour lesquelles la membrane plasmique présynaptique est fortement enrichie en lipides polyinsaturés, en particulier lorsque l'expansion de la tige est l'étape limitant la vitesse de la réaction de fusion.
4. La formation de la tige et son expansion, l'ouverture et l'expansion du pore de fusion seraient donc contrôlés par la courbure des lipides (Synergistic actions of v-SNARE transmembrane domains and membrane-curvature modifying lipids in neurotransmitter release 2020).
- Étant donné la capacité du feuillet interne à s'étirer, la barrière résultante contre l'élargissement de la tige est extrêmement sensible à la distribution des lipides entre les feuillets externe et interne (Expansion of the fusion stalk and its implication for biological membrane fusion 2014).
- Même de légères variations de la répartition des lipides entre les monocouches (quelques % seulement) donneront déjà lieu à une tension inter-feuillets importante, qui peut modifier la barrière d'énergie libre de plusieurs dizaines de kBT
Déshydratation partielle des lipides
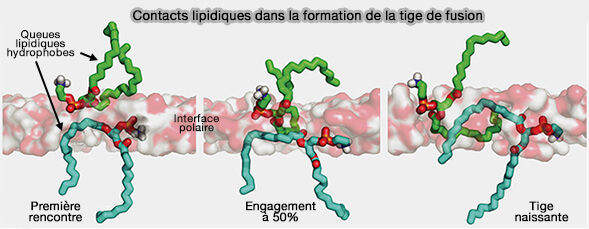
Le lien et le motif justamembranaire (JM) de la synaptobrévine 2/VAMP2 provoqueraient une déshydratation partielle du feuillet externe membranaire qui modifierait l'orientation des groupes de tête lipidique ( rôles du lien et du domaine transmembranaire (TMD) dans la formation du complexe trans-SNARE).
rôles du lien et du domaine transmembranaire (TMD) dans la formation du complexe trans-SNARE).
(Figure : vetopsy.fr d'après Kasson et coll)
1. La présence de phospholipides chargés négativement ainsi que la courbure de la membrane influencent également fortement la probabilité de protrusion de leurs chaînes acyles (Atomic-Resolution Simulations Predict a Transition State for Vesicle Fusion Defined by Contact of a Few Lipid Tails 2010).
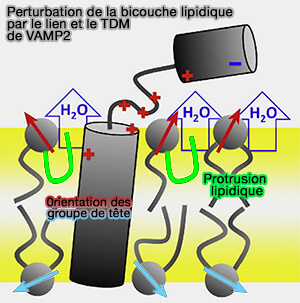
(Figure : vetopsy.fr d'après Han et coll)
L'état de transition pour la formation de la tige se produit lorsqu'une paire de queues lipidiques entre en contact à travers la couche d'interface polaire. Dans les simulations, le contact entre une seule paire de queues hydrophobes serait suffisant pour nucléer la formation de tiges de fusion.
2. La perturbation de la bicouche lipidique par la la synaptobrévine 2/VAMP2 :
- favorise la protrusion lipidique,
- influence l'orientation du groupe de tête (flèches rouges et jaunes).

Le feuillet interne d'une membrane plasmique typique de mammifère est plus fusiogène que le feuillet externe d'environ 50kJ/mol.
Remarque : les protéines volumineuses de liaison aux SNARE abaissent la barrière d'énergie libre contre l'élargissement de la tige.

La figure ci-dessous représente l'évolution des vésicules du stade d'ancrage à la fusion complète ( lipides et formation du pore de fusion).
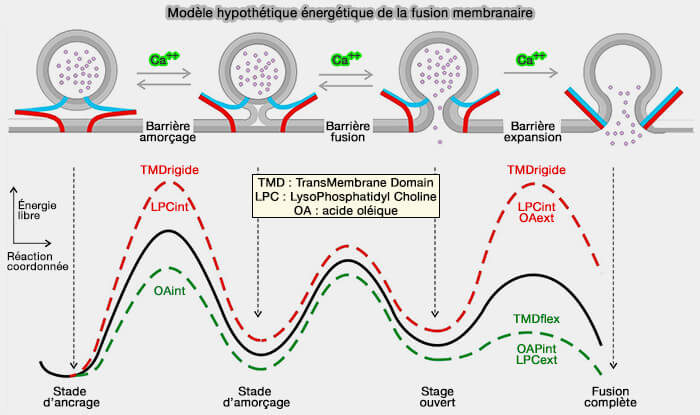
(Figure : vetopsy.fr d'après Dhara et coll)
Synaptotagmine et tige de fusion
1. La synaptotagmine-1 est un capteur calcique Ca++ de faible affinité qui déclenche la fusion synchrone des vésicules.
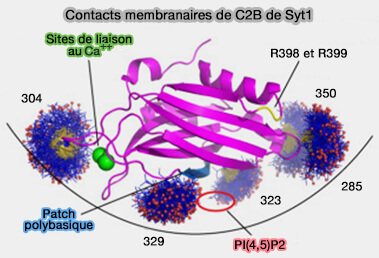
(Figure : vetopsy.fr d'après Nyenhuis et coll)
Elles contiennent deux domaines C2, C2A et C2B qui coopèrent dans la liaison membranaire, le domaine C2B étant principalement responsable du processus de fusion membranaire en raison de son patch polybasique 321KRLKKKKTTIKK332 et de son apex d'arginine ( synaptotagmine et formation du pore de fusion).
Les synaptotagmines sont longuement étudiées dans des chapitres spéciaux.
2. Un mécanisme maître/serviteur entre deux domaines C2B identiques contrôle la formation de la tige de fusion, d'une manière indépendante du calcium(Synaptotagmin-1 C2B domains cooperatively stabilize the fusion stalk via a master-servant mechanism 2022).
- Le domaine maître montre de fortes interactions avec PI(4,5)P2 ou PIP2 de la membrane plasmique au niveau de l'apex d'arginine et du patch polybasique.
- Dans le même temps, le serviteur coopère de manière analogue avec le maître pour réduire le travail total pour former la tige de fusion.
Les mutations T328E et T329E dans les domaines maître et serviteur perturbe le mécanisme coopératif, augmentant considérablement l'énergie libre nécessaire pour induire la tige de fusion.
- Elles ont des effets négligeables sur les interactions du domaine maître avec PIP2.
- Elles perturbent le comportement du domaine serviteur, incapable de maintenir ses interactions PIP2, ni par son patch polybasique, ni par sa paire d'arginines, et au final, perd sa capacité à assister le maître dans la formation de la tige de fusion.
Formation du pore de fusion
Biologie cellulaire et moléculaireConstituants de la celluleTransport membranaireTransports sans mouvements membranairesTransports passifsTransports actifsTransports avec mouvements membranaires : trafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireCanaux ioniquesTransporteursUniportsPompesCo-transporteursMoteurs moléculairesVoies de signalisation