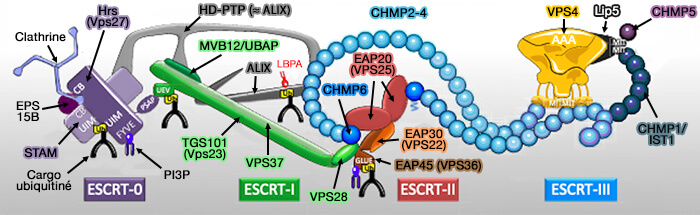
Le complexe ESCRT (Endosomal Sorting Complex Required for Transport ou complexe de tri endosomal requis pour le transport) est composé de 5 complexes :
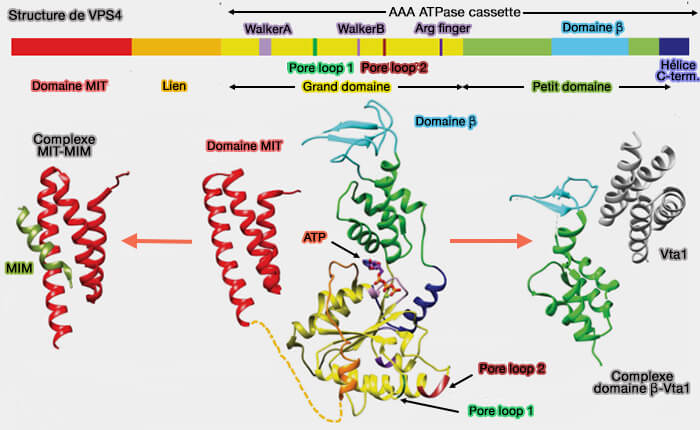
L'extrémité C-terminale de ESCRT-III recrute plusieurs molécules effectrices dont la protéine VPS4 (AAA ATPase vacuolaire de tri 4) qui a un rôle essentiel
:
1. VPS4 fait partie de la grande famille des AAA+ ATPases qui a été classée en plusieurs clades dont le clade méiotique avec les enzymes spastine, katanine et fidgétine.
Toutes ces enzymes désassemblent des structures protéiques polymères spécifiques :
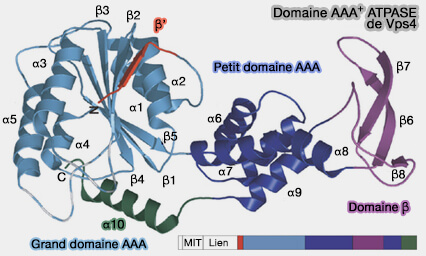
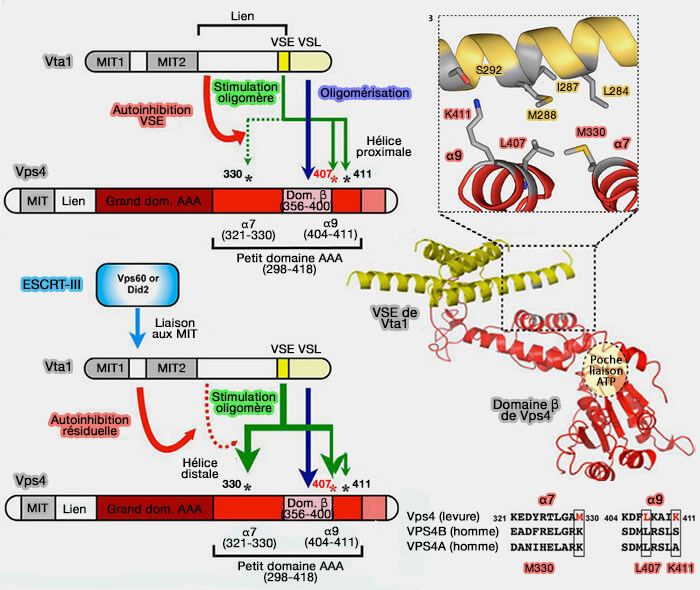
Le petit domaine ATPase comprend quatre hélices, suivie d'une hélice C-terminale qui le compacte contre le grand domaine. Le domaine β émane d'une boucle interne du petit domaine ATPase et interagit pour avec LIP5/Vta1.
2. L'hélice à cinq sous-unités est droite avec une translation de ~6,3Å et une rotation de 60° entre les sous-unités, i.e. correspondant à la symétrie du peptide ESCRT-III lié, qui adopte une conformation de feuillet β (β-Strand).
Ces spirales autour de l'axe hélicoïdal se lient dans une rainure formée par les cinq sous-unités hélicoïdales VPS4. Les peptides de substrat successifs, DEIVNKVL dans l'article, se lient de la même manière aux interfaces successives entre les sous-unités VPS4.
La chaîne latérale du premier résidu de chaque dipeptide se lie dans une poche de classe I, tandis que la chaîne latérale du deuxième résidu de chaque dipeptide se lie dans une poche de classe II. et ces poches sont répétées à chacune des quatre interfaces des sous-unités VPS4 hélicoïdales.
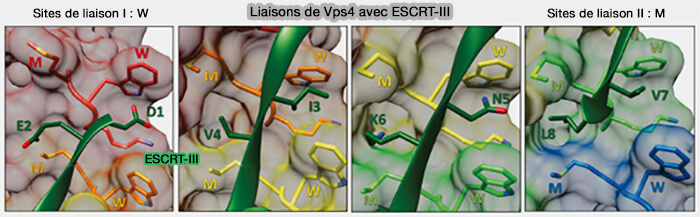
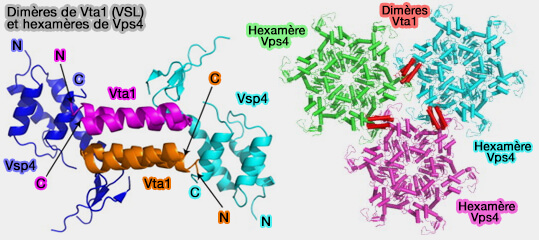
Liaisons Vps4-ESCRT-III
(Figure : vetopsy.fr d'après Han
et Hill)
3. Les sites de liaison :
de classe I sont principalement formées par le résidu de tryptophane (W) de la boucle de pore 1 qui forme une encoche hydrophobe qui apparaît capable d'accueillir une grande variété de chaînes latérales d'acides aminés (premier résidu) entre deux sous-unités VPS4 adjacentes,
de classe II contiennent une méthionine (M) hydrophobe flanquée par la boucle de pore 2 qui peut servir de poche, avec la sous-unité adjacente, à une grande variété d'acides aminés.
La liaison est largement indépendante de la séquence pour la translocation du substrat de protéines ESCRT-III.
Recyclage de ESCRT-III
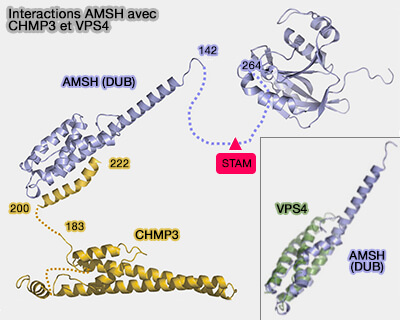
Interactions entre Vps4 et ESCRT-III
Interactions MIT/MIM des CHMP
Le recyclage induit par Vps4 dépend de l'interaction du domaine MIT (Microtubule Interacting and Trafficking molecule domain) avec les motifs d'interaction MIM (Mit Interaction Motif) des sous-unités ESCRT-III.
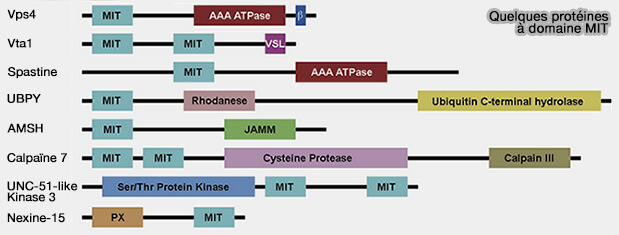
Quelques protéines à domaine MIT
(Figure : vetopsy.fr d'après Huxley
et Yang)
Les domaines MIT se composent de trois faisceaux d'hélices qui interagissent avec des partenaires protéiques via plusieurs interfaces.
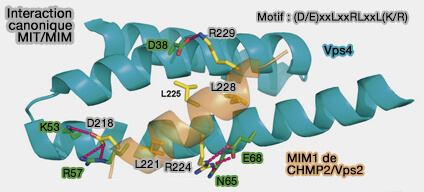
Interaction canonique MIT/MIM
(Figure : vetopsy.fr d'après Hurley et Yang)
1. L'interaction canonique d'ESCRT-III avec le domaine MIT de Vps4 se produit à travers MIM1 des sous-unités :
Le MIM1 consiste en une seule hélice qui interagit avec les hélices α2 et 3 du domaine MIT de Vps4 via plusieurs résidus Leu clés de MIM1 (D/E)xxLxxRLxxL(K/R).
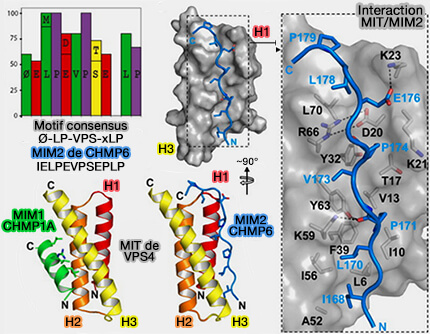
MIT de Vps4 avec MIM2 de CHMP6
(Figure : vetopsy.fr d'après Kieffer
et coll)
CHMP3 n'a pas de MIM interagissant avec VPS4, mais CHMP3 et la plupart des autres sous-unités ESCRT-III ont des séquences qui se lient à d'autres protéines à domaines MIT.
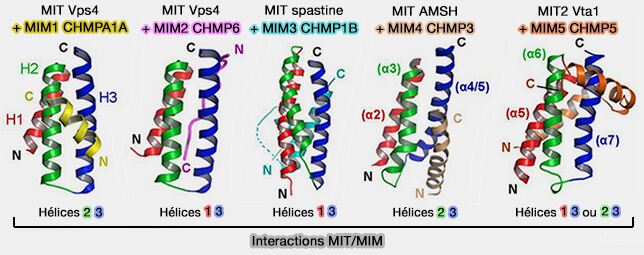
Interactions MIT/MIM
(Figure : vetopsy.fr d'après Yang et coll)
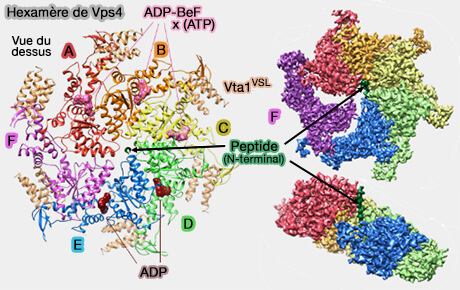
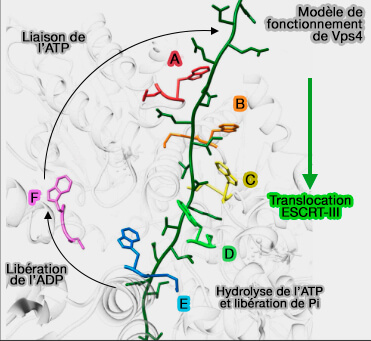
1. Dans le modèle de l'hexamérisation asymétrique décrit plus haut, il y a plusieurs modèles, Vps4 procède par transition entre les configurations des sous-unités autour de l'anneau hexamérique, i.e. transition de la configuration des sous-unités F, A, B, C, D, E aux configurations de sous-unités A, B, C, D, E, F.
Modèle de fonctionnement de Vps4
(Figure : vetopsy.fr d'après Han
et Hill)
Vps4 " marche " le long des ESCRT-III par l'action des sous-unités qui :
dénudent l'extrémité arrière de l'hélice formée par les cinq sous-unités, i.e. la sous-unité E,
passent par la conformation transitoire, i.e. déplacement de la sous-unité F,
permettent à l'extrémité de l'hélice, i.e. la sous-unité A, de se lier au dipeptide suivant dans la séquence ESCRT-III.
Conformément à ce modèle, le grand domaine ATPase de la sous-unité F est déplacé hors de l'axe de l'hélice, dégagé du peptide ESCRT-III, et n'a que
des contacts mineurs avec les grands domaines de sous-unités voisines Vps4. La sous-unité F est maintenue dans le complexe Vps4 principalement par des contacts plus périphériques des petits domaines d'ATPase.
2. Une caractéristique intéressante de ce modèle est que chaque sous-unité Vps4 n'a besoin d'adopter qu'une conformation et que son dipeptide ESCRT-III associé passe à travers le pore de la position de la sous-unité A à la position de la sous-unité E.
Il semblerait que l'interface des sous-unités EF soit plus ouvert que les autres, ce qui favoriserait l'hydrolysation de l'ATP à l'interface DE, i.e. provoquent la déstabilisation des interactions et une conformation EF plus lâche.
Vps4 n'aurait besoin de déplier qu'environ la moitié des sous-unités d'un filament afin d'en démonter l'intégralité.
3. Quand l'affinité des CHMP pour Vps4 est faible (CHMP4) ou négligeable (CHMP3), on n'a pas encore explicité comment se fait le recyclage.
Les sous-unités à affinité forte (CHMP1, CHMP2, CHMP6 et IST1) aident les faibles à se dissocier.
Lip5/Vta1 peut aider ce désassemblage.
Une fois le recyclage des sous-unités à affinité forte effectué, tout s'effondre.



 Rôles de la spastine avec ESCRT-III dans les endosomes de recyclage).
Rôles de la spastine avec ESCRT-III dans les endosomes de recyclage).