Embryologie du système nerveux : induction neurale
Gradients
Citation
ÂŦ Commander, conduire et guider, non par dures ni violentes contraintes, mais par molles et douces inductions et persuasions. Âŧ
Jacques Amyot
Documentation web
- Biologie du dÃĐveloppement
- ÂŦ De l'oeuf à la grenouille Âŧ Les Etapes du DÃĐveloppement d'un Amphibien : Le xenope (Xenopus laevis)
- Biologie du dÃĐveloppement animal
- DÃĐveloppement embryonnaire des VertÃĐbrÃĐs
- Gastrulation
- Embryologie molÃĐculaire
- Embryologie humaine
- Embryologie gÃĐnÃĐrale
- Biologie du dÃĐveloppement
Lâinduction neurale se fait selon 2 gradients :
- un gradient cÃĐphalo-caudal,
- un gradient dorso-ventral.
(Figure : © vetopsy.fr d'aprÃĻs Fred the Oyster)
Lâinduction neurale se fait selon 3 gradients.
Gradient cÃĐphalo-caudal
Le gradient cÃĐphalo-caudal, c'est-Ã -dire, la multiplication cellulaire plus rostrale que caudale, explique l'allongement du germe, sphÃĐrique jusqu'alors. La plaque neurale est :
- étroite en position caudale, à l'origine de la future moelle épinière,
- élargie en position antérieure, en forme de raquette, à l'origine du futur encÃĐphale.
![]()
Le gÃĻne Nodal (cf. plus bas) a un rÃīle primordial dans la mise en place de l'axe antÃĐro-postÃĐrieur, ainsi que de la latÃĐralitÃĐ droit-gauche.
Cette induction est aussi diffÃĐrente selon l'axe antÃĐro-postÃĐrieur du tube neural (polaritÃĐ antÃĐro-postÃĐrieure).
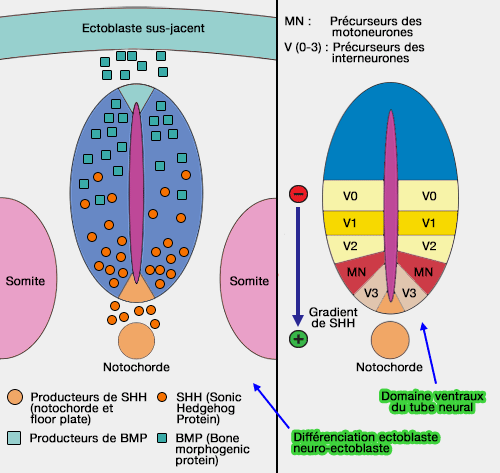
Gradient dorso-ventral
Le gradient dorso-ventral s'exprime par le ligand SHH (Sonic Hedgedog), issu de la notochorde, et de la plaque prÃĐchordale pour le tube neural antÃĐrieur (polaritÃĐ dorso-ventrale : cf. figure ci-dessus).
Les concentrations en SHH varient selon un gradient :
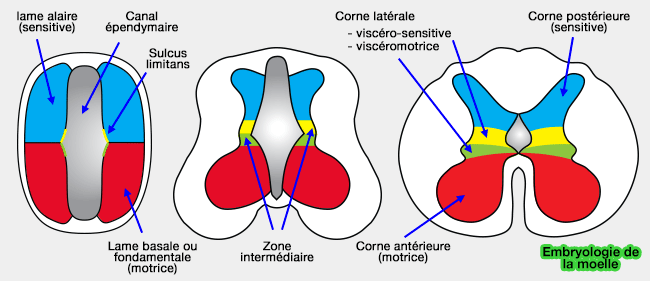
- les moins ÃĐlevÃĐes se situent au niveau de la partie dorsale du tube neural : les lames alaires, dorsales, sont sous le contrÃīle des TGF-β (dorsaline, activine, BMP4) et de Wnts qui facilitent l'expression de gÃĻnes comme PAX 3 et 7.
- les concentrations maximales se retrouvent au niveau de la plaque du plancher (floor plate), c'est-Ã -dire les cellules les plus ventrales du tube neural. Ces cellules expriment alors ÃĐgalement la SHH (voie autocrine) et ce mÃĐcanisme rÃĐprime alors des gÃĻnes comme PAX 3, 7 et 6 (" gÃĻnes dorsalisants ") et permet le dÃĐveloppement des motoneurones.
(Figure : © vetopsy.fr)
Ce mÃĐcanisme permet, via la concentration mais aussi le temps d'exposition à la SHH, l'expression de divers facteurs de transcription qui aboutit à la diffÃĐrenciation des cellules neuronales, en particulier les neurones moteurs, sensitifs et les interneurones.
La SHH est donc impliquÃĐe dans de nombreux processus.
1. Formation de structures paires comme les hÃĐmisphÃĻres cÃĐrÃĐbraux.
Une anomalie de la SHH en rÃĐgion crÃĒniale provoque la formation d'un cerveau impair et mÃĐdian indiffÃĐrenciÃĐ.
2. DÃĐtermination des cÃītÃĐs droit et gauche de l'embryon (latÃĐralitÃĐ droite-gauche).
![]()
Le gÃĻne Nodal, surexprimÃĐ Ã gauche, induit la latÃĐralitÃĐ dÃĻs la formation du noeud et de la ligne primitive grÃĒce à la SHH.
(Figure : © vetopsy.fr d'aprÃĻs Rthistle)
{kind=link}
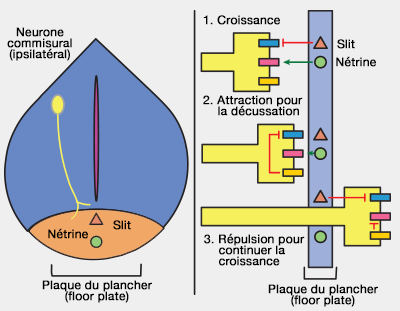
3. La dÃĐcussation ou non de certains neurones.
Les neurones controlatÃĐraux se dÃĐveloppent tout d'abord dans la rÃĐgion dorsale et se dÃĐplacent vers le plancher pour traverser la ligne mÃĐdiane embryonnaire en formant les commissures. Ce guidage est ÃĐgalement sous le contrÃīle des nÃĐtrines et du slit à effet opposÃĐ.
- Les nÃĐtrines attirent les neurones commissuraux vers la ligne mÃĐdiane en se liant au rÃĐcepteur de l'axone DCC de maniÃĻre chimiotactique.
- Le slit les repoussent en interagissant sur les rÃĐcepteurs transmembranaires Roundabout (Robo1 et Robo2). Cette voie de signalisation slit-robo inhibe la chimiotaxie de la voie nÃĐtrine/DCC. En outre, pendant la migration des neurones commissuraux vers la plaque du plancher, la prÃĐsence de Robo3 inhibe l'expression de Robo1 et 2.
Formation de l'encÃĐphale
SystÃĻme nerveux![]() Embryologie du systÃĻme nerveux : vue d'ensemble
Embryologie du systÃĻme nerveux : vue d'ensemble![]() Induction neurale
Induction neurale
- Houillon Ch. - Embryologie - Hermann, Paris, 184 p., 1979
- Kahle W. - Anatomie, Tome 3 Système nerveux et organes des sens - Flammarion Médecine-Sciences, 423 p., 2007
- Marieb E. N. - Anatomie et physiologie humaines - De Boeck Université, Saint-Laurent, 1054 p., 1993
- Rosenzweig M.R., Leiman A.L., Breedlove S.M. - Psychobiologie - DeBoeck Université, Bruxelles, 849 p., 1998
- Purves D., Augustine G.J., Fitzpatrick D., Katz L.C., Lamantia A-S, McNamara J.O., Williams S.M. - Neurosciences - De Boeck, 800 p., 2003
- Kolb B., Whishaw I. - Cerveau et comportement - De Boeck, 646 p., 2002
- Bear M.F., Connors B.W., Paradiso M.A. - Neurosciences : à la découverte du cerveau - Masson-Williams, 654 p, 1997
- Gazzaniga, Ivry, Mangun - Neurosciences cognitives - De Boeck, 585 p., 2001